

A direct descending pathway informing locomotor networks about tactile sensor movement
- PMID: 25740535
- PMCID: PMC6605569
- DOI: 10.1523/JNEUROSCI.3350-14.2015
A direct descending pathway informing locomotor networks about tactile sensor movement
Abstract
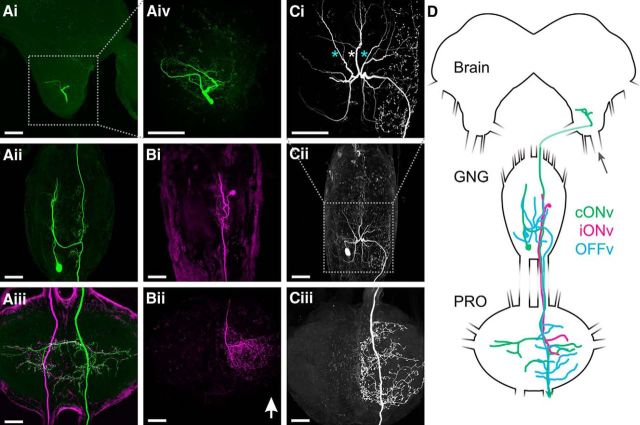
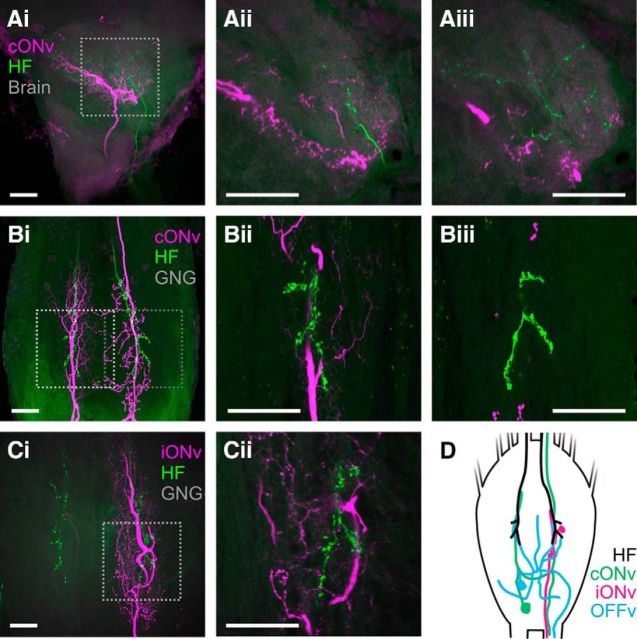
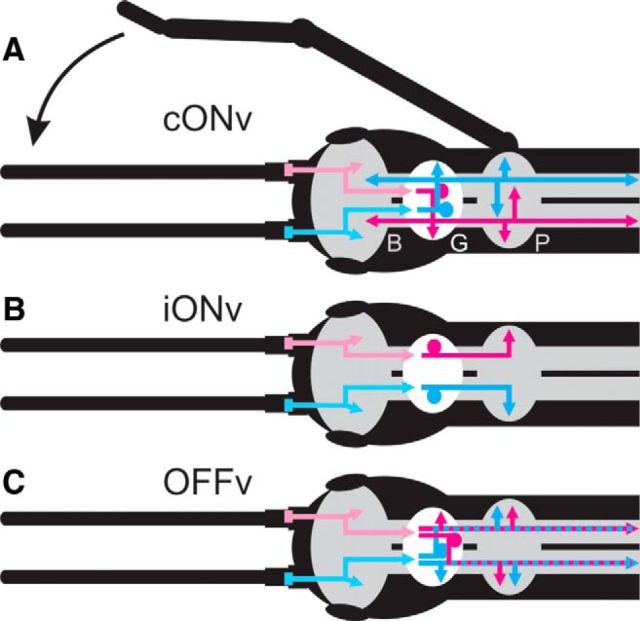
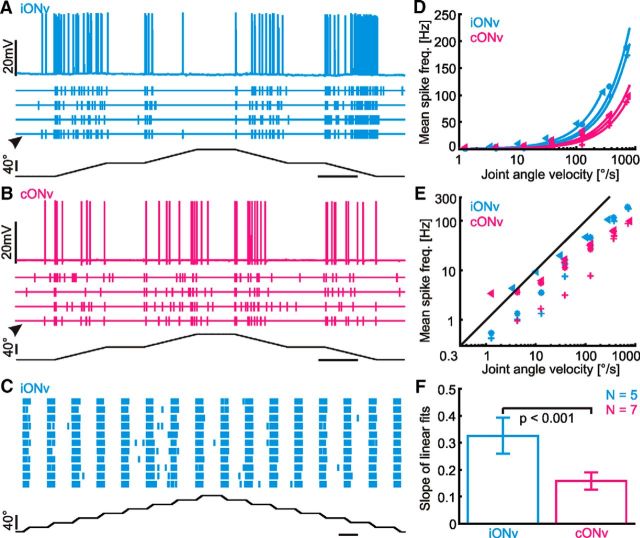
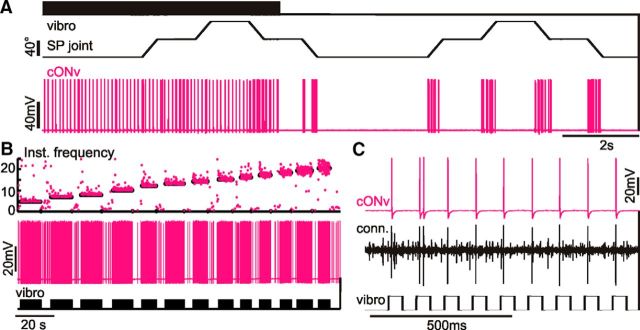
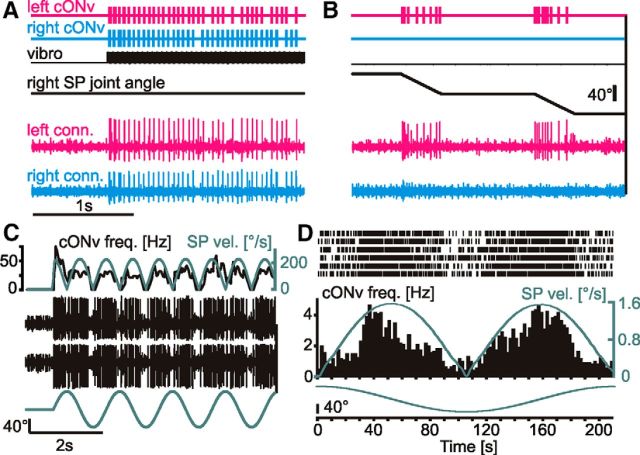
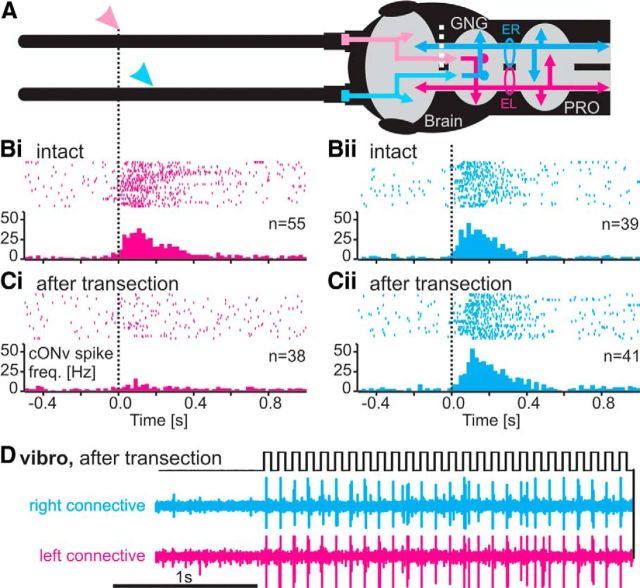
Much like visually impaired humans use a white-cane, nocturnal insects and mammals use antennae or whiskers for near-range orientation. Stick insects, for example, rely heavily on antennal tactile cues to find footholds and detect obstacles. Antennal contacts can even induce aimed reaching movements. Because tactile sensors are essentially one-dimensional, they must be moved to probe the surrounding space. Sensor movement is thus an essential cue for tactile sensing, which needs to be integrated by thoracic networks for generating appropriate adaptive leg movements. Based on single and double recordings, we describe a descending neural pathway comprising three identified ON- and OFF-type neurons that convey complementary, unambiguous, and short-latency information about antennal movement to thoracic networks in the stick insect. The neurons are sensitive to the velocity of antennal movements across the entire range covered by natural movements, regardless of movement direction and joint angle. Intriguingly, none of them originates from the brain. Instead, they descend from the gnathal ganglion and receive input from antennal mechanoreceptors in this lower region of the CNS. From there, they convey information about antennal movement to the thorax. One of the descending neurons, which is additionally sensitive to substrate vibration, feeds this information back to the brain via an ascending branch. We conclude that descending interneurons with complementary tuning characteristics, gains, input and output regions convey detailed information about antennal movement to thoracic networks. This pathway bypasses higher processing centers in the brain and thus constitutes a shortcut between tactile sensors on the head and the thorax.
Keywords: adaptive locomotion; descending control; identified neurons; invertebrates; tactile sensing.
Copyright © 2015 the authors 0270-6474/15/354081-11$15.00/0.
Figures

References
-
- Alstermark B, Lindström S, Lundberg A, Sybirska E. Integration in descending motor pathways controlling the forelimb in the cat: 8. Ascending projection to the lateral reticular nucleus from C3–C4 propriospinal also projecting to forelimb motoneurones. Exp Brain Res. 1981;42:282–298. - PubMed
-
- Bacon JP, Strausfeld NJ. The dipteran giant fibre pathway: neurons and signals. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 1986;158:529–548. doi: 10.1007/BF00603798. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources