

Physico-chemical requirements and kinetics of membrane fusion of flavivirus-like particles
- PMID: 25740960
- PMCID: PMC4635454
- DOI: 10.1099/vir.0.000113
Physico-chemical requirements and kinetics of membrane fusion of flavivirus-like particles
Abstract
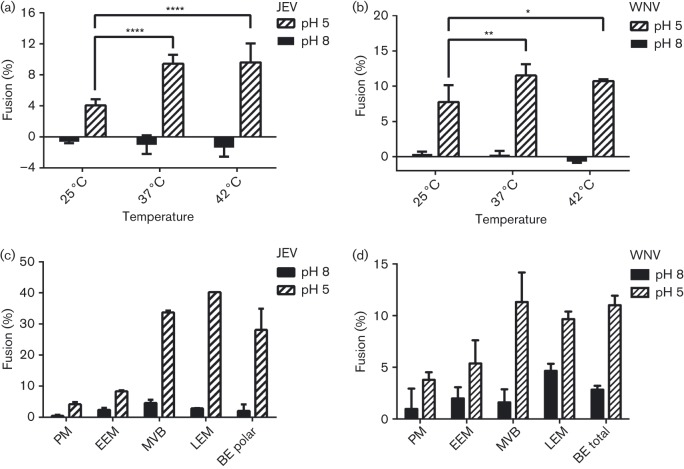
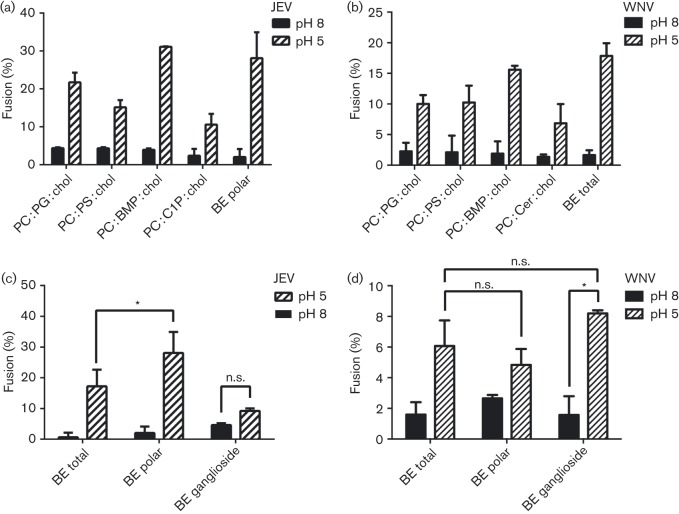
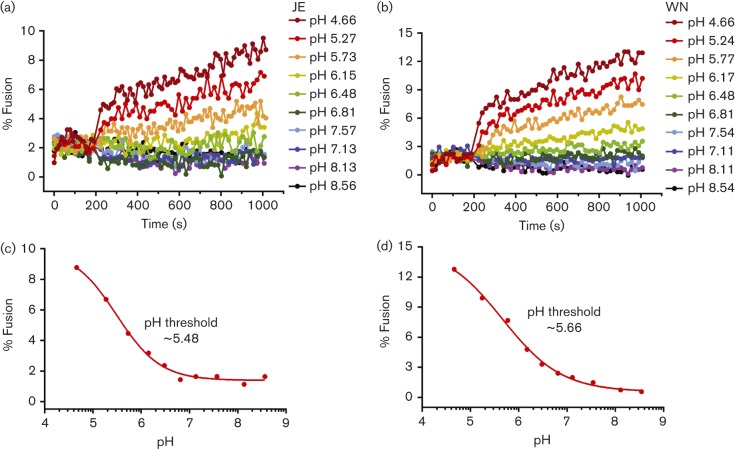
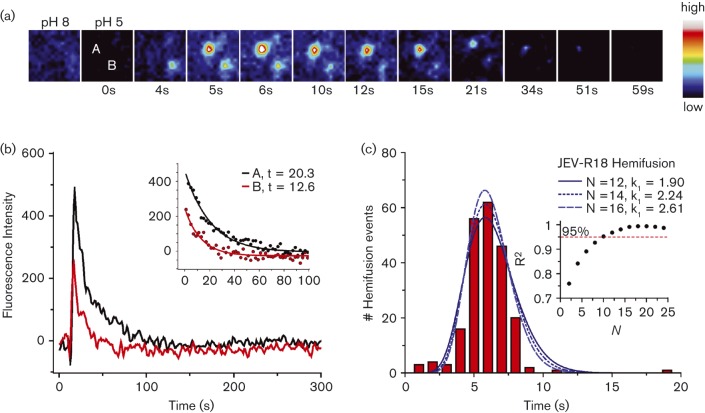
Flaviviruses deliver their RNA genome into the host-cell cytoplasm by fusing their lipid envelope with a cellular membrane. Expression of the flavivirus pre-membrane and envelope glycoprotein genes in the absence of other viral genes results in the spontaneous assembly and secretion of virus-like particles (VLPs) with membrane fusion activity. Here, we examined the physico-chemical requirements for membrane fusion of VLPs from West Nile and Japanese encephalitis viruses. In a bulk fusion assay, optimal hemifusion (or lipid mixing) efficiencies were observed at 37 °C. Fusion efficiency increased with decreasing pH; half-maximal hemifusion was attained at pH 5.6. The anionic lipids bis(monoacylglycero)phosphate and phosphatidylinositol-3-phosphate, when present in the target membrane, significantly enhanced fusion efficiency, consistent with the emerging model that flaviviruses fuse with intermediate-to-late endosomal compartments, where these lipids are most abundant. In a single-particle fusion assay, VLPs catalysed membrane hemifusion, tracked as lipid mixing with the cellular membrane, on a timescale of 7-20 s after acidification. Lipid mixing kinetics suggest that hemifusion is a kinetically complex, multistep process.
Figures
References
-
- Bajjalieh S. M., Martin T. F., Floor E. ( 1989. ). Synaptic vesicle ceramide kinase. A calcium-stimulated lipid kinase that co-purifies with brain synaptic vesicles. J Biol Chem 264, 14354–14360. - PubMed
-
- Das S., Chakraborty S., Basu A. ( 2010. ). Critical role of lipid rafts in virus entry and activation of phosphoinositide 3′ kinase/Akt signaling during early stages of Japanese encephalitis virus infection in neural stem/progenitor cells. J Neurochem 115, 537–549. 10.1111/j.1471-4159.2010.06951.x - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous