

Role of EXT1 and Glycosaminoglycans in the Early Stage of Filovirus Entry
- PMID: 25741008
- PMCID: PMC4442511
- DOI: 10.1128/JVI.03689-14
Role of EXT1 and Glycosaminoglycans in the Early Stage of Filovirus Entry
Abstract
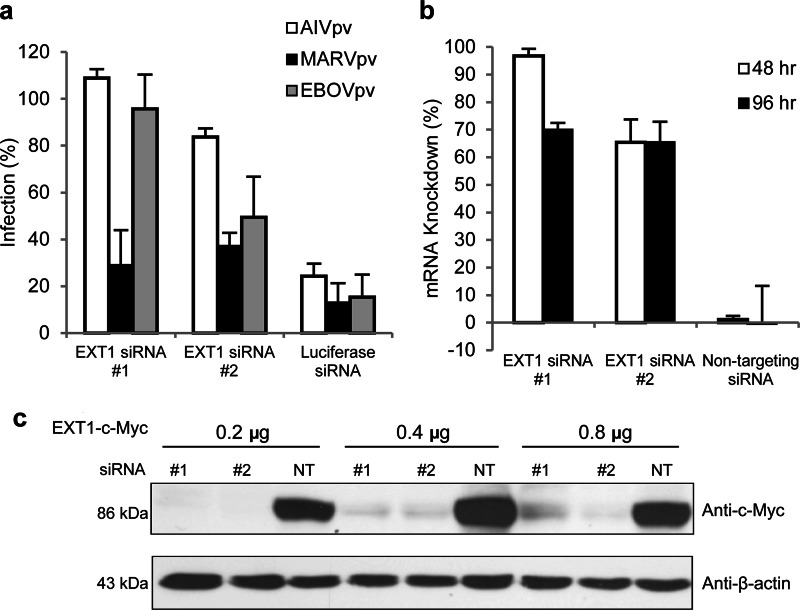
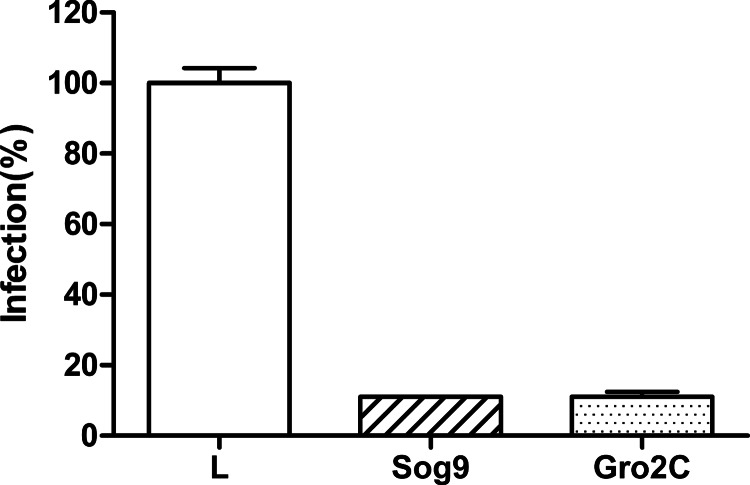
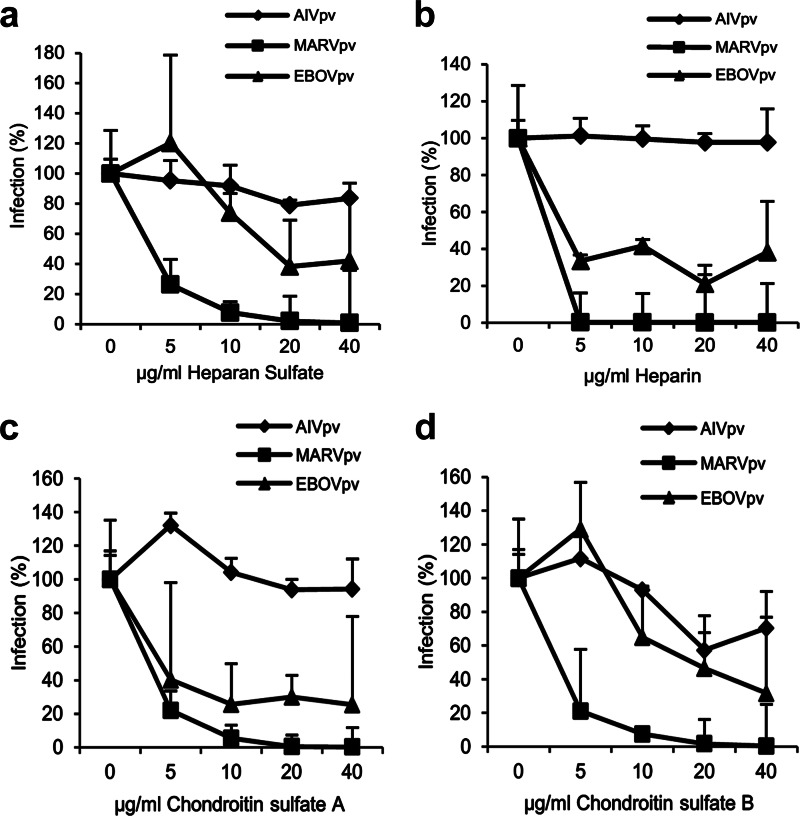
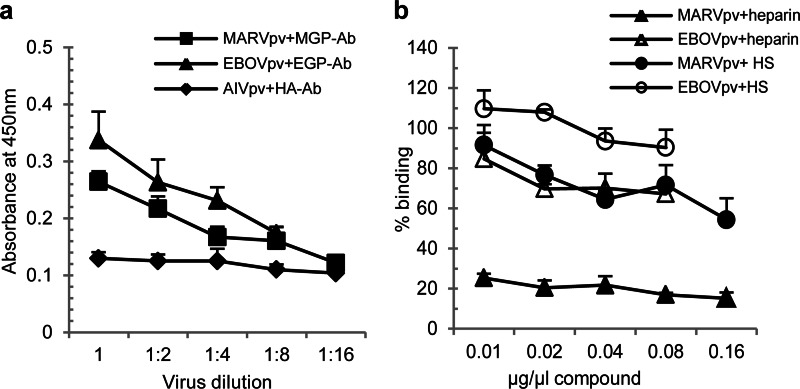
Filoviruses, including both Ebola virus (EBOV) and Marburg virus (MARV), can infect humans and other animals, causing hemorrhagic fever with a high mortality rate. Entry of these viruses into the host is mediated by a single filoviral glycoprotein (GP). GP is composed of two subunits: GP1, which is responsible for attachment and binding to receptor(s) on susceptible cells, and GP2, which mediates viral and cell membrane fusion. Although numerous host factors have been implicated in the entry process, the initial attachment receptor(s) has not been well defined. In this report, we demonstrate that exostosin 1 (EXT1), which is involved in biosynthesis of heparan sulfate (HS), plays a role in filovirus entry. Expression knockdown of EXT1 by small interfering RNAs (siRNAs) impairs GP-mediated pseudoviral entry and that of infectious EBOV and MARV in tissue cultured cells. Furthermore, HS, heparin, and other related glycosaminoglycans (GAGs), to different extents, can bind to and block GP-mediated viral entry and that of infectious filoviruses. These results strongly suggest that HS and other related GAGs are attachment receptors that are utilized by filoviruses for entry and infection. These GAGs may have therapeutic potential in treating EBOV- and MARV-infected patients.
Importance: Infection by Ebola virus and Marburg virus can cause severe illness in humans, with a high mortality rate, and currently there is no FDA-approved vaccine or therapeutic treatment available. The ongoing 2014 outbreak in West Africa underscores a lack of our understanding in the infection and pathogenesis of these viruses and the urgency of drug discovery and development. In this study, we provide several pieces of evidence that demonstrate that heparan sulfate and other closely related glycosaminoglycans are the molecules that are used by filoviruses for initial attachment. Furthermore, we demonstrate that these glycosaminoglycans can block entry of and infection by filoviruses. Thus, this work provides mechanistic insights on the early step of filoviral infection and suggests a possible therapeutic option for diseases caused by filovirus infection.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Inhibition of Ebola and Marburg Virus Entry by G Protein-Coupled Receptor Antagonists.J Virol. 2015 Oct;89(19):9932-8. doi: 10.1128/JVI.01337-15. Epub 2015 Jul 22. J Virol. 2015. PMID: 26202243 Free PMC article.
-
Molecular virulence determinants of human-pathogenic filoviruses.Adv Virus Res. 2025;121:1-29. doi: 10.1016/bs.aivir.2025.03.003. Epub 2025 Apr 2. Adv Virus Res. 2025. PMID: 40379380 Review.
-
Interaction between TIM-1 and NPC1 Is Important for Cellular Entry of Ebola Virus.J Virol. 2015 Jun;89(12):6481-93. doi: 10.1128/JVI.03156-14. Epub 2015 Apr 8. J Virol. 2015. PMID: 25855742 Free PMC article.
-
A biaryl sulfonamide derivative as a novel inhibitor of filovirus infection.Antiviral Res. 2020 Nov;183:104932. doi: 10.1016/j.antiviral.2020.104932. Epub 2020 Sep 15. Antiviral Res. 2020. PMID: 32946918 Free PMC article.
-
Filovirus entry: a novelty in the viral fusion world.Viruses. 2012 Feb;4(2):258-75. doi: 10.3390/v4020258. Epub 2012 Feb 7. Viruses. 2012. PMID: 22470835 Free PMC article. Review.
Cited by
-
Inhibition of Ebola and Marburg Virus Entry by G Protein-Coupled Receptor Antagonists.J Virol. 2015 Oct;89(19):9932-8. doi: 10.1128/JVI.01337-15. Epub 2015 Jul 22. J Virol. 2015. PMID: 26202243 Free PMC article.
-
Identification of a coumarin-based antihistamine-like small molecule as an anti-filoviral entry inhibitor.Antiviral Res. 2017 Sep;145:24-32. doi: 10.1016/j.antiviral.2017.06.015. Epub 2017 Jun 20. Antiviral Res. 2017. PMID: 28645623 Free PMC article.
-
Screening and Reverse-Engineering of Estrogen Receptor Ligands as Potent Pan-Filovirus Inhibitors.J Med Chem. 2020 Oct 8;63(19):11085-11099. doi: 10.1021/acs.jmedchem.0c01001. Epub 2020 Sep 22. J Med Chem. 2020. PMID: 32886512 Free PMC article.
-
Polyphenylene carboxymethylene (PPCM) in vitro antiviral efficacy against Ebola virus in the context of a sexually transmitted infection.Antiviral Res. 2019 Oct;170:104567. doi: 10.1016/j.antiviral.2019.104567. Epub 2019 Jul 24. Antiviral Res. 2019. PMID: 31351092 Free PMC article.
-
Structure of the Ebola virus glycoprotein spike within the virion envelope at 11 Å resolution.Sci Rep. 2017 Apr 11;7:46374. doi: 10.1038/srep46374. Sci Rep. 2017. PMID: 28397863 Free PMC article.
References
-
- Feldmann F, Sanchez A, Geisbert JB. 2013. Filoviridae: Marburg and Ebola viruses, p 923–956. In Knipe DM, Howley PM (ed), Fields virology, 6th ed, vol 2 Wolters Kluwer, Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Qiu X, Wong G, Audet J, Bello A, Fernando L, Alimonti JB, Fausther-Bovendo H, Wei H, Aviles J, Hiatt E, Johnson A, Morton J, Swope K, Bohorov O, Bohorova N, Goodman C, Kim D, Pauly MH, Velasco J, Pettitt J, Olinger GG, Whaley K, Xu B, Strong JE, Zeitlin L, Kobinger GP. 2014. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 514:47–53. doi:10.1038/nature13777. - DOI - PMC - PubMed
-
- Warren TK, Wells J, Panchal RG, Stuthman KS, Garza NL, Van Tongeren SA, Dong L, Retterer CJ, Eaton BP, Pegoraro G, Honnold S, Bantia S, Kotian P, Chen X, Taubenheim BR, Welch LS, Minning DM, Babu YS, Sheridan WP, Bavari S. 2014. Protection against filovirus diseases by a novel broad-spectrum nucleoside analogue BCX4430. Nature 508:402–405. doi:10.1038/nature13027. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
