

doi: 10.1128/JVI.03652-14.
Epub 2015 Mar 4.
Ebola virus VP35 interaction with dynein LC8 regulates viral RNA synthesis
Affiliations
- PMID: 25741013
- PMCID: PMC4403485
- DOI: 10.1128/JVI.03652-14
Item in Clipboard
Ebola virus VP35 interaction with dynein LC8 regulates viral RNA synthesis
J Virol.
2015 May.
Abstract
Ebola virus VP35 inhibits alpha/beta interferon production and functions as a viral polymerase cofactor. Previously, the 8-kDa cytoplasmic dynein light chain (LC8) was demonstrated to interact with VP35, but the functional consequences were unclear. Here we demonstrate that the interaction is direct and of high affinity and that binding stabilizes the VP35 N-terminal oligomerization domain and enhances viral RNA synthesis. Mutational analysis demonstrates that VP35 interaction is required for the functional effects of LC8.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
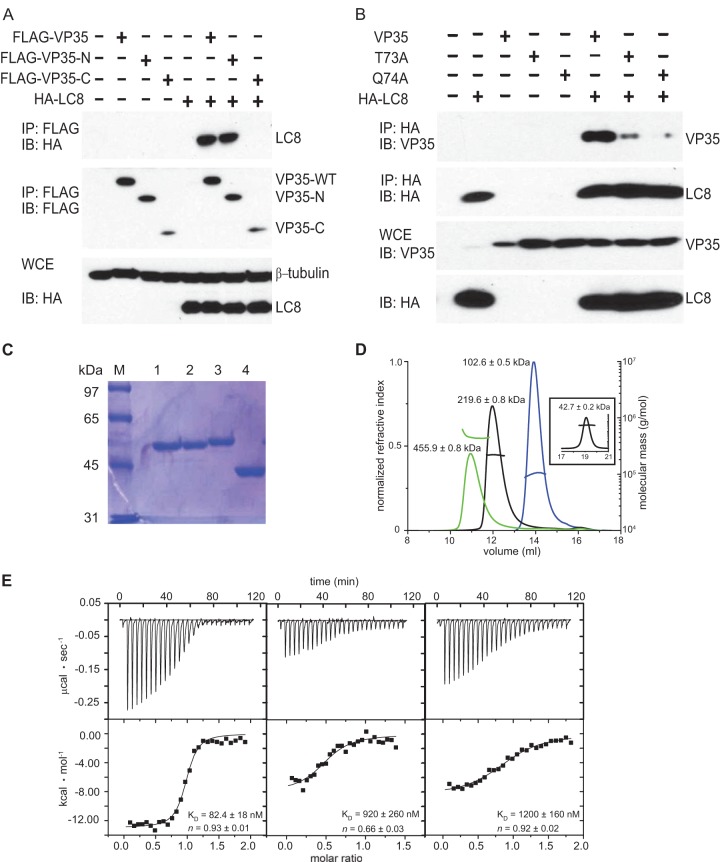
LC8 interacts with EBOV VP35 protein. (A) The N terminus of VP35 is sufficient to interact with LC8. A co-IP experiment was performed with FLAG-tagged WT VP35, VP35-N (containing amino acids [aa] 1 to 218), VP35-C (aa 219 to 340), and HA-tagged LC8 (HA-LC8). VP35 proteins were pulled down by anti-FLAG antibody, and protein expression was detected by Western blotting (immunoblotting [IB]). WCE, whole-cell extracts. (B) VP35 mediates the interaction with LC8 via an LC8 binding consensus motif (K/SXTQT). VP35 mutants (the T73A and Q74A mutants) were utilized in a co-IP experiment with HA-tagged LC8. Protein expression was detected by Western blotting with anti-VP35 and anti-HA antibodies. (C) SDS-PAGE gel of MBP-VP35 50–150 Q74A (lane 1), MBP-VP35 50–150 (lane 2), MBP-LC8 (lane 3), and MBP (lane 4) used in SEC-MALS and ITC experiments. (D) VP35 forms a 1:1 complex with LC8. Elution profiles of MBP-LC8 (blue), MBP-VP35 50–150 (black), MBP-VP35 50–150/MBP-LC8 complex (green), and MBP (inset) from a Superdex 200 column were obtained and analyzed by SEC-MALS. The theoretical monomeric molecular masses for MBP-VP35 50–150 and MBP-LC8 are 55.2 and 54.4 kDa, respectively. (E) ITC data for MBP-LC8 binding to MBP-WT VP35 50–150 (left), mutant MBP-VP35 50–150 Q74A (center), and mutant MBP-VP35 50–150 Q72A Q74A (right). Representative raw data and binding isotherms are shown, and the reported values for KD and n are averages from at least two independent experiments.
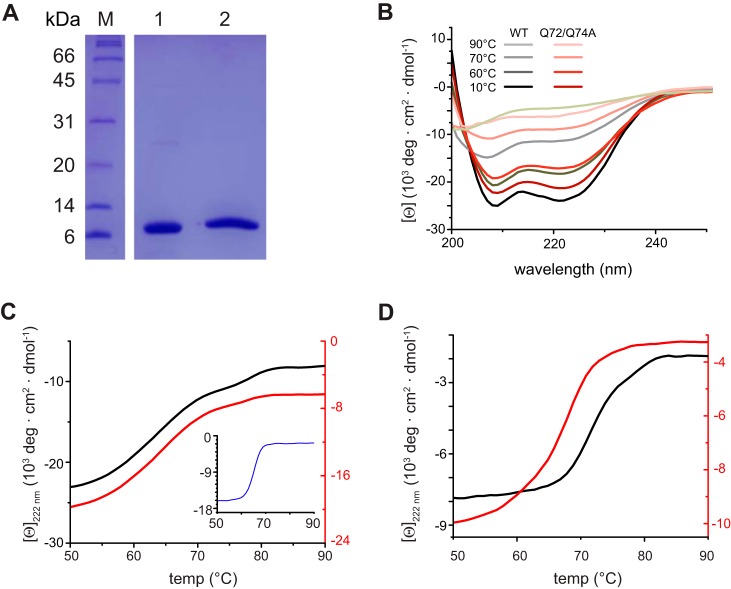
LC8 binding enhances the stability of the N terminus of VP35. (A) SDS-PAGE gel of VP35 50–150 (lane 1) and LC8 (lane 2) used in circular dichroism (CD) experiments. Lane M, molecular mass markers. (B) CD wavelength scans for WT VP35 50–150 (black lines) and the VP35 50–150 Q72A Q74A mutant (red lines) at the temperatures indicated. Mean residue ellipticity is in 103 degrees · cm2 · dmol−1. (C) Thermal denaturation (Tm) of WT VP35 50–150 (black, left axis; Tm = 63.8 ± 0.1°C), the VP35 50–150 Q72A Q74A mutant (red, right axis; Tm = 63.8 ± 1°C), and LC8 (inset, blue; Tm = 66.3 ± 2°C) monitored at 222 nm. (D) Thermal denaturation of the WT VP35 50–150/LC8 complex (black, left axis; Tm = 70.8 ± 1°C) and the VP35 50–150 Q72A Q74A/LC8 mutant complex (red, right axis; Tm = 66.4 ± 2°C) monitored at 222 nm. Reported Tms are averages of results from at least two independent experiments.
LC8 enhances EBOV RNA synthesis in a dose-dependent manner. (A) Increasing amounts of HA-tagged LC8 plasmid (50, 500 ng) were cotransfected with the plasmids required to reconstitute the EBOV RNA polymerase complex (L, NP, VP35, VP30) along with a plasmid encoding the Renilla luciferase minigenome RNA and a firefly luciferase expression plasmid, which served as a control for transfection efficiency. Relative activity was determined by normalizing Renilla luciferase activity to firefly luciferase activity. The error bars indicate standard deviations from three independent replicates. *, P = 0.04; **, P = 0.03, as determined by Student's t test. The Western blot shows expression of β-tubulin, VP35, and LC8 (anti-HA antibody). (B) A minigenome experiment similar to that described for panel A was performed except that WT VP35 or the indicated VP35 mutants were used, and the LC8 plasmid amount was kept constant at 500 ng. The error bars indicate standard deviations from three independent replicates. The Western blot shows the expression of VP35, LC8, and β-tubulin, detected with anti-VP35, anti-HA, and anti-β-tubulin antibodies, respectively. *, P = 0.01; **, P = 0.006, as determined by Student's t test.
LC8 expression enhances EBOV transcription. (A) A minigenome assay was performed as described for Fig. 2A, except that a replication-deficient minigenome construct was used. *, P = 0.01; **, P = 0.0045, as determined by Student's t test. (B) A minigenome experiment similar to that described for panel A was performed except that WT VP35 or the indicated VP35 mutants were included and the LC8 plasmid amount was kept constant at 500 ng. The error bars indicate standard deviations from three independent replicates. The Western blot shows the expression of VP35 and LC8, detected with anti-VP35 and anti-HA antibodies, respectively. *, P = 0.007; **, P = 0.002, as determined by Student's t test.
Similar articles
-
Virus and host interactions critical for filoviral RNA synthesis as therapeutic targets.Antiviral Res. 2019 Feb;162:90-100. doi: 10.1016/j.antiviral.2018.12.006. Epub 2018 Dec 11. Antiviral Res. 2019. PMID: 30550800 Free PMC article. Review.
-
Crystal structure of human LC8 bound to a peptide from Ebola virus VP35.J Microbiol. 2021 Apr;59(4):410-416. doi: 10.1007/s12275-021-0641-7. Epub 2021 Feb 25. J Microbiol. 2021. PMID: 33630249
-
The Host E3-Ubiquitin Ligase TRIM6 Ubiquitinates the Ebola Virus VP35 Protein and Promotes Virus Replication.J Virol. 2017 Aug 24;91(18):e00833-17. doi: 10.1128/JVI.00833-17. Print 2017 Sep 15. J Virol. 2017. PMID: 28679761 Free PMC article.
-
Basic residues within the ebolavirus VP35 protein are required for its viral polymerase cofactor function.J Virol. 2010 Oct;84(20):10581-91. doi: 10.1128/JVI.00925-10. Epub 2010 Aug 4. J Virol. 2010. PMID: 20686031 Free PMC article.
-
DRBP76 associates with Ebola virus VP35 and suppresses viral polymerase function.J Infect Dis. 2011 Nov;204 Suppl 3(Suppl 3):S911-8. doi: 10.1093/infdis/jir343. J Infect Dis. 2011. PMID: 21987769 Free PMC article.
Cited by
-
Virus and host interactions critical for filoviral RNA synthesis as therapeutic targets.Antiviral Res. 2019 Feb;162:90-100. doi: 10.1016/j.antiviral.2018.12.006. Epub 2018 Dec 11. Antiviral Res. 2019. PMID: 30550800 Free PMC article. Review.
-
Current status of small molecule drug development for Ebola virus and other filoviruses.Curr Opin Virol. 2019 Apr;35:42-56. doi: 10.1016/j.coviro.2019.03.001. Epub 2019 Apr 16. Curr Opin Virol. 2019. PMID: 31003196 Free PMC article. Review.
-
Crystal Structure of the Marburg Virus VP35 Oligomerization Domain.J Virol. 2017 Jan 3;91(2):e01085-16. doi: 10.1128/JVI.01085-16. Print 2017 Jan 15. J Virol. 2017. PMID: 27847355 Free PMC article.
-
Dynein Light Chain LC8 Is Required for RNA Polymerase I-Mediated Transcription in Trypanosoma brucei, Facilitating Assembly and Promoter Binding of Class I Transcription Factor A.Mol Cell Biol. 2015 Oct 12;36(1):95-107. doi: 10.1128/MCB.00705-15. Print 2016 Jan 1. Mol Cell Biol. 2015. PMID: 26459761 Free PMC article.
-
Single-Cell Profiling of Ebola Virus Disease In Vivo Reveals Viral and Host Dynamics.Cell. 2020 Nov 25;183(5):1383-1401.e19. doi: 10.1016/j.cell.2020.10.002. Epub 2020 Nov 6. Cell. 2020. PMID: 33159858 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI081914/AI/NIAID NIH HHS/United States
- T32-CA09547-37/CA/NCI NIH HHS/United States
- R01 AI107056/AI/NIAID NIH HHS/United States
- U19 AI070489/AI/NIAID NIH HHS/United States
- R01AI081914/AI/NIAID NIH HHS/United States
- T32 AI007163/AI/NIAID NIH HHS/United States
- R01AI059536/AI/NIAID NIH HHS/United States
- R01 AI059536/AI/NIAID NIH HHS/United States
- U19 AI109945/AI/NIAID NIH HHS/United States
- P01 AI120943/AI/NIAID NIH HHS/United States
- U19AI109945/AI/NIAID NIH HHS/United States
- R01AI107056/AI/NIAID NIH HHS/United States
- U19 AI109664/AI/NIAID NIH HHS/United States
- U19AI109664/AI/NIAID NIH HHS/United States
- T32 CA009547/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
