

T-cell intracellular antigens function as tumor suppressor genes
- PMID: 25741594
- PMCID: PMC4385921
- DOI: 10.1038/cddis.2015.43
T-cell intracellular antigens function as tumor suppressor genes
Abstract
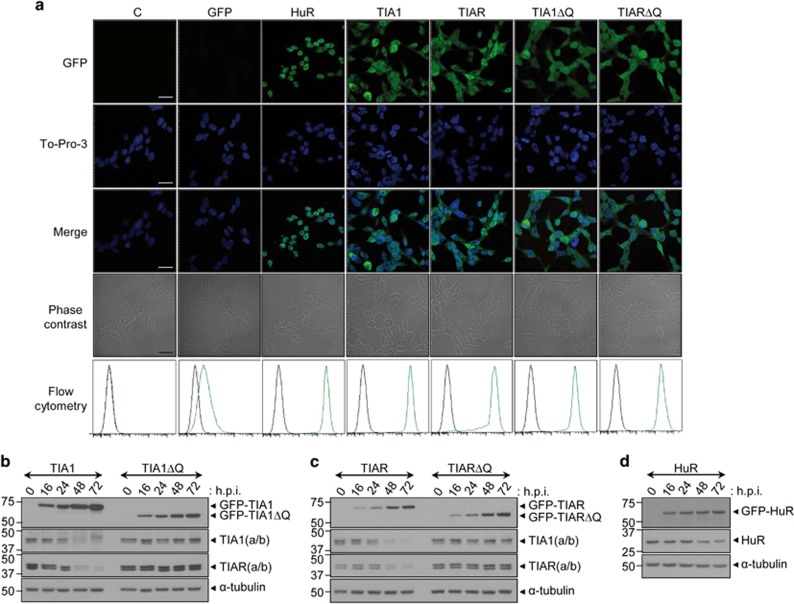
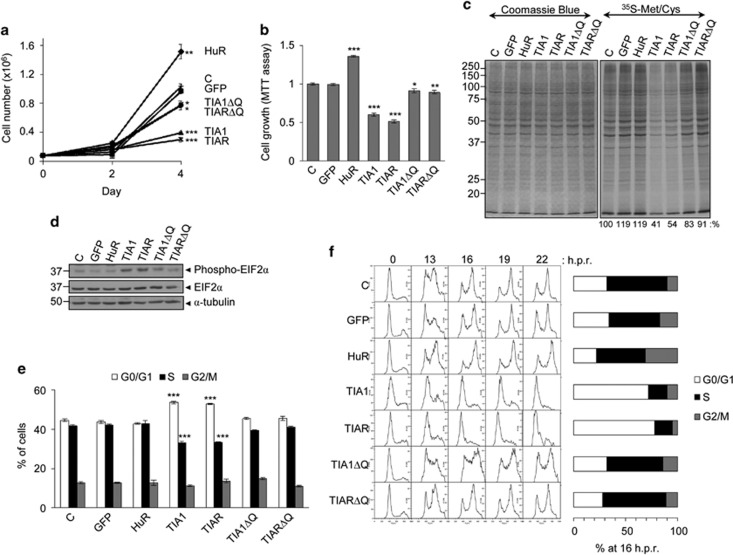
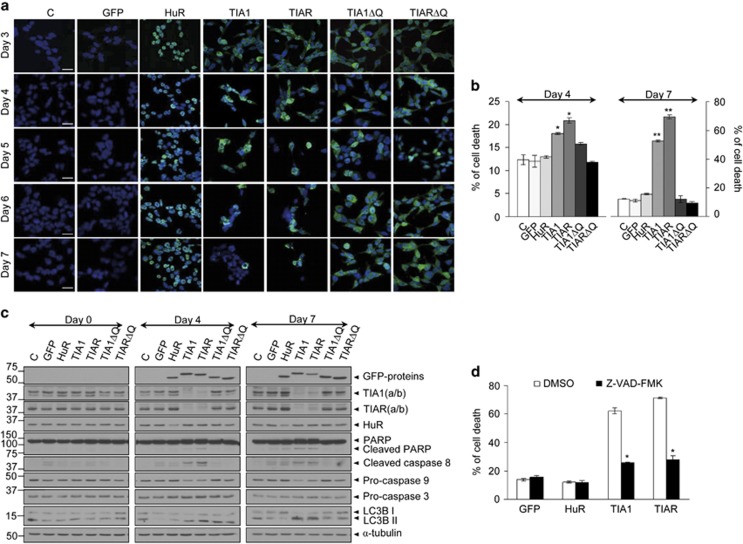
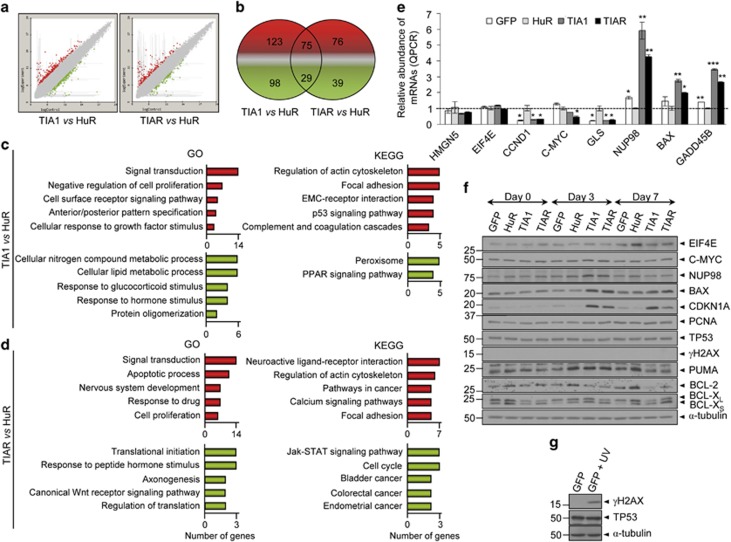
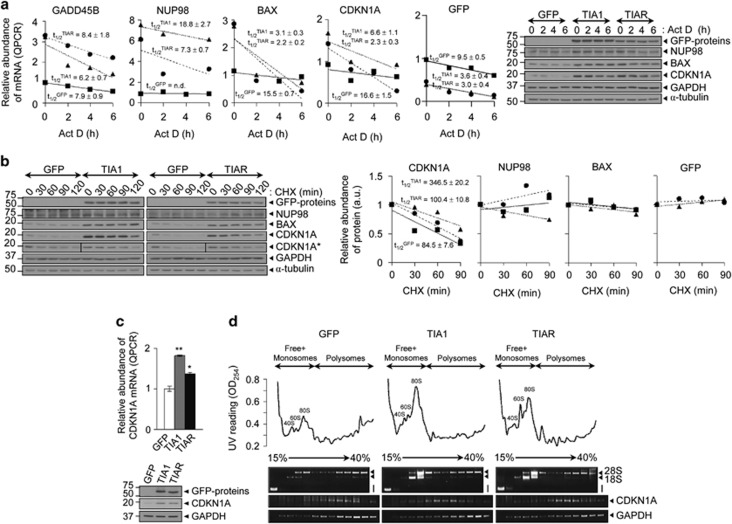
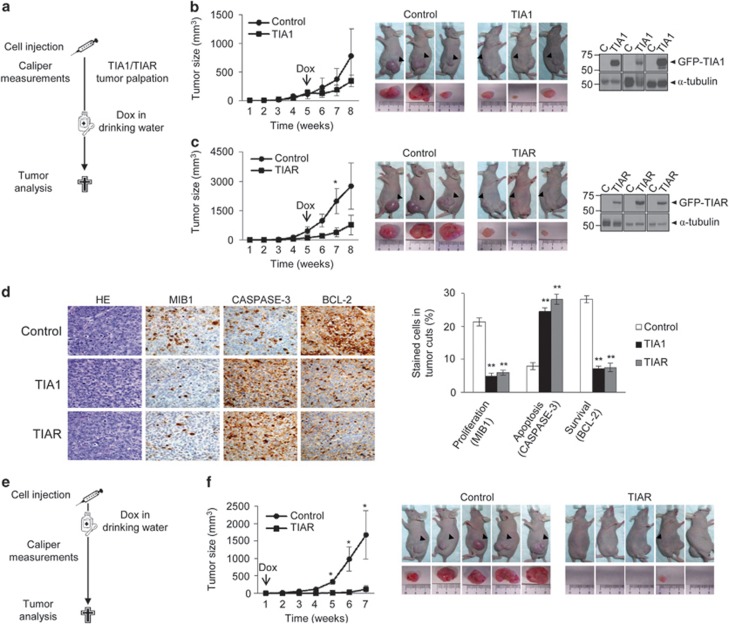
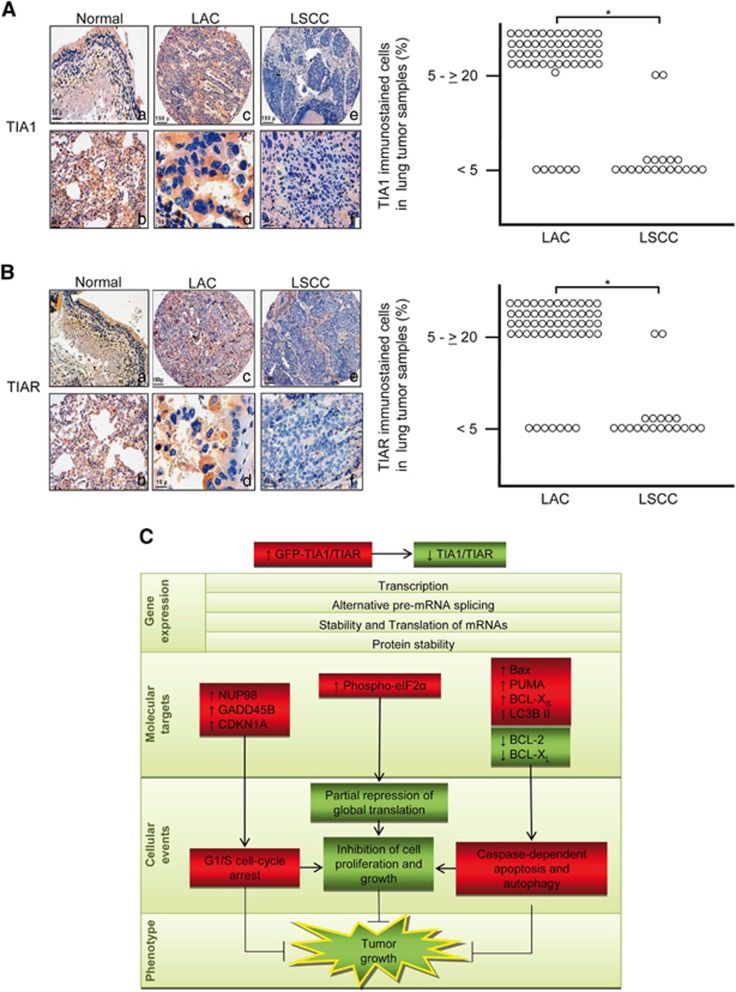
Knockdown of T-cell intracellular antigens TIA1 and TIAR in transformed cells triggers cell proliferation and tumor growth. Using a tetracycline-inducible system, we report here that an increased expression of TIA1 or TIAR in 293 cells results in reduced rates of cell proliferation. Ectopic expression of these proteins abolish endogenous TIA1 and TIAR levels via the regulation of splicing of their pre-mRNAs, and partially represses global translation in a phospho-eukaryotic initiation factor 2 alpha-dependent manner. This is accompanied by cell cycle arrest at G1/S and cell death through caspase-dependent apoptosis and autophagy. Genome-wide profiling illustrates a selective upregulation of p53 signaling pathway-related genes. Nude mice injected with doxycycline-inducible cells expressing TIA1 or TIAR retard, or even inhibit, growth of xenotumors. Remarkably, low expressions of TIA1 and TIAR correlate with poor prognosis in patients with lung squamous cell carcinoma. These findings strongly support the concept that TIA proteins act as tumor suppressor genes.
Figures
Similar articles
-
T-Cell Intracellular Antigen 1-Like Protein in Physiology and Pathology.Int J Mol Sci. 2022 Jul 16;23(14):7836. doi: 10.3390/ijms23147836. Int J Mol Sci. 2022. PMID: 35887183 Free PMC article. Review.
-
T-cell intracellular antigens in health and disease.Cell Cycle. 2015;14(13):2033-43. doi: 10.1080/15384101.2015.1053668. Cell Cycle. 2015. PMID: 26036275 Free PMC article. Review.
-
Knockdown of T-cell intracellular antigens triggers cell proliferation, invasion and tumour growth.Biochem J. 2011 Apr 15;435(2):337-44. doi: 10.1042/BJ20101030. Biochem J. 2011. PMID: 21284605
-
TIAR and TIA-1 mRNA-binding proteins co-aggregate under conditions of rapid oxygen decline and extreme hypoxia and suppress the HIF-1α pathway.J Mol Cell Biol. 2010 Dec;2(6):345-56. doi: 10.1093/jmcb/mjq032. Epub 2010 Oct 27. J Mol Cell Biol. 2010. PMID: 20980400
-
T-cell intracellular antigen (TIA)-proteins deficiency in murine embryonic fibroblasts alters cell cycle progression and induces autophagy.PLoS One. 2013 Sep 24;8(9):e75127. doi: 10.1371/journal.pone.0075127. eCollection 2013. PLoS One. 2013. PMID: 24086455 Free PMC article.
Cited by
-
T-Cell Intracellular Antigen 1-Like Protein in Physiology and Pathology.Int J Mol Sci. 2022 Jul 16;23(14):7836. doi: 10.3390/ijms23147836. Int J Mol Sci. 2022. PMID: 35887183 Free PMC article. Review.
-
LncRNA MT1JP functions as a tumor suppressor by interacting with TIAR to modulate the p53 pathway.Oncotarget. 2016 Mar 29;7(13):15787-800. doi: 10.18632/oncotarget.7487. Oncotarget. 2016. PMID: 26909858 Free PMC article.
-
The emerging role of alternative splicing in senescence and aging.Aging Cell. 2017 Oct;16(5):918-933. doi: 10.1111/acel.12646. Epub 2017 Jul 13. Aging Cell. 2017. PMID: 28703423 Free PMC article. Review.
-
Stress granules dynamics: benefits in cancer.BMB Rep. 2022 Dec;55(12):577-586. doi: 10.5483/BMBRep.2022.55.12.141. BMB Rep. 2022. PMID: 36330685 Free PMC article. Review.
-
Application of RNA processing factors for predicting clinical outcomes in colon cancer.Front Genet. 2022 Sep 23;13:979001. doi: 10.3389/fgene.2022.979001. eCollection 2022. Front Genet. 2022. PMID: 36212157 Free PMC article.
References
-
- Tian Q, Streuli M, Saito H, Schlossman SF, Anderson P. A polyadenylate binding protein localized to the granules of cytolytic lymphocytes induces DNA fragmentation in target cells. Cell. 1991;67:629–639. - PubMed
-
- McAlinden A, Liang L, Mukudai Y, Imamura T, Sandell LJ. Nuclear protein TIA1 regulates COL2A1 alternative splicing and interacts with precursor mRNA and genomic DNA. J Biol Chem. 2007;282:24444–24454. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
