

Osteoblast ablation reduces normal long-term hematopoietic stem cell self-renewal but accelerates leukemia development
- PMID: 25742698
- PMCID: PMC4408292
- DOI: 10.1182/blood-2014-06-582924
Osteoblast ablation reduces normal long-term hematopoietic stem cell self-renewal but accelerates leukemia development
Abstract
Hematopoietic stem cells (HSCs) reside in regulatory niches in the bone marrow (BM). Although HSC niches have been extensively characterized, the role of endosteal osteoblasts (OBs) in HSC regulation requires further clarification, and the role of OBs in regulating leukemic stem cells (LSCs) is not well studied. We used an OB visualization and ablation mouse model to study the role of OBs in regulating normal HSCs and chronic myelogenous leukemia (CML) LSCs. OB ablation resulted in increase in cells with a LSK Flt3(-)CD150(+)CD48(-) long-term HSC (LTHSC) phenotype but reduction of a more highly selected LSK Flt3(-)CD34(-)CD49b(-)CD229(-) LTHSC subpopulation. LTHSCs from OB-ablated mice demonstrated loss of quiescence and reduced long-term engraftment and self-renewal capacity. Ablation of OB in a transgenic CML mouse model resulted in accelerated leukemia development with reduced survival compared with control mice. The notch ligand Jagged-1 was overexpressed on CML OBs. Normal and CML LTHSCs cultured with Jagged-1 demonstrated reduced cell cycling, consistent with a possible role for loss of Jagged-1 signals in altered HSC and LSC function after OB ablation. These studies support an important role for OBs in regulating quiescence and self-renewal of LTHSCs and a previously unrecognized role in modulating leukemia development in CML.
© 2015 by The American Society of Hematology.
Figures
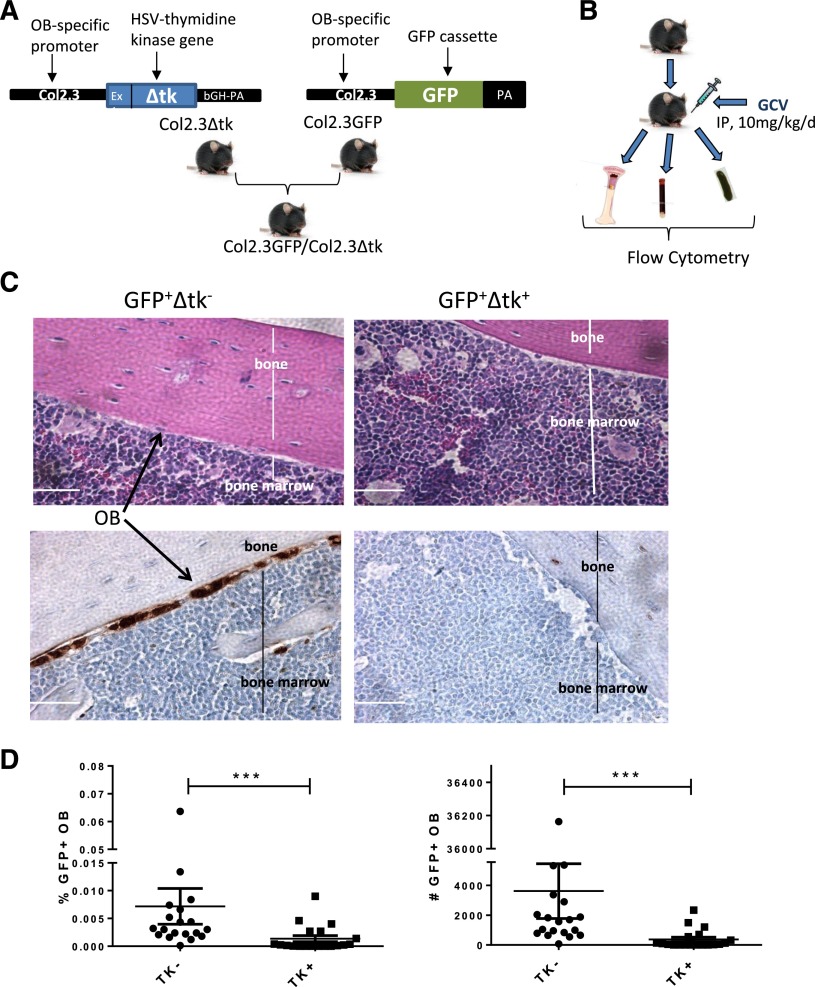


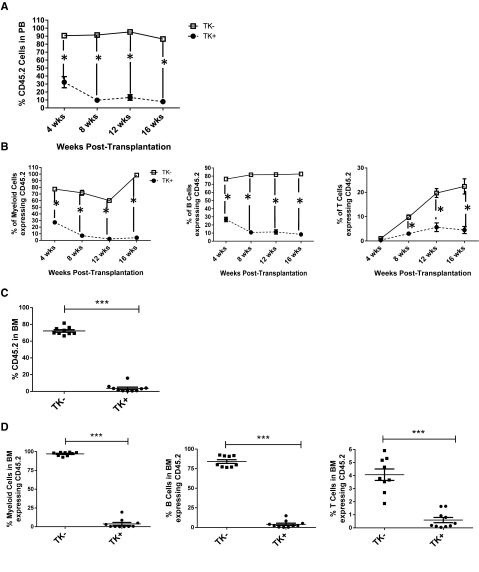
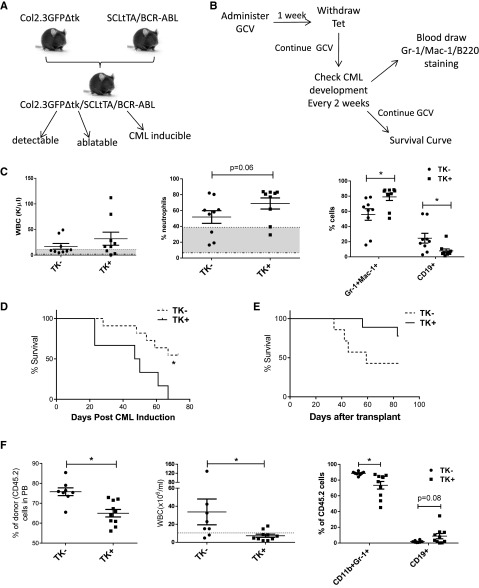
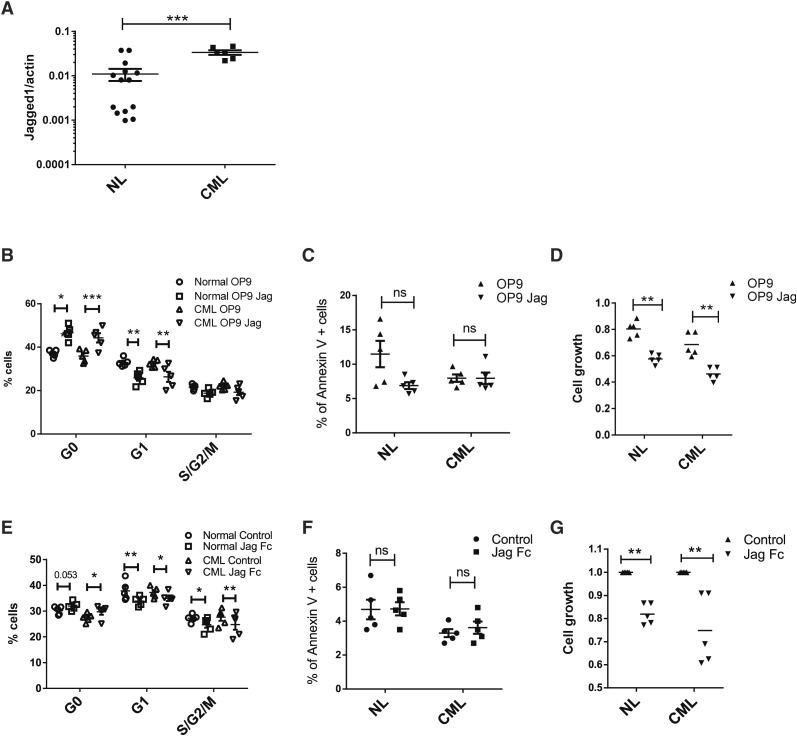
Comment in
-
Osteoblast ablation burns out functional stem cells.Blood. 2015 Apr 23;125(17):2590-1. doi: 10.1182/blood-2015-03-633651. Blood. 2015. PMID: 25907901
References
-
- Arai F, Suda T. Maintenance of quiescent hematopoietic stem cells in the osteoblastic niche. Ann N Y Acad Sci. 2007;1106:41–53. - PubMed
-
- Visnjic D, Kalajzic Z, Rowe DW, Katavic V, Lorenzo J, Aguila HL. Hematopoiesis is severely altered in mice with an induced osteoblast deficiency. Blood. 2004;103(9):3258–3264. - PubMed
-
- Calvi LM, Adams GB, Weibrecht KW, et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature. 2003;425(6960):841–846. - PubMed
-
- Guezguez B, Campbell CJ, Boyd AL, et al. Regional localization within the bone marrow influences the functional capacity of human HSCs. Cell Stem Cell. 2013;13(2):175–189. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
