

IL4 receptor α mediates enhanced glucose and glutamine metabolism to support breast cancer growth
- PMID: 25746764
- PMCID: PMC4380623
- DOI: 10.1016/j.bbamcr.2015.02.020
IL4 receptor α mediates enhanced glucose and glutamine metabolism to support breast cancer growth
Abstract
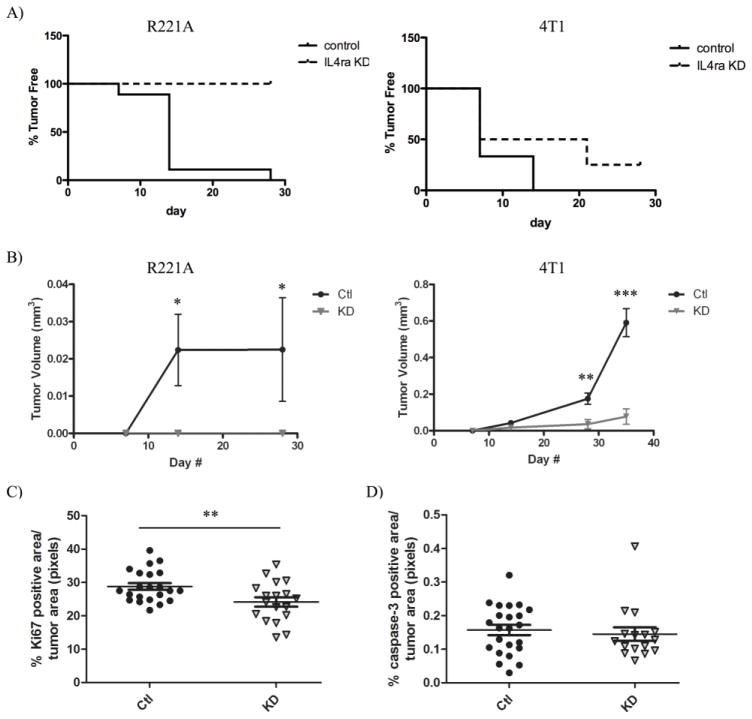
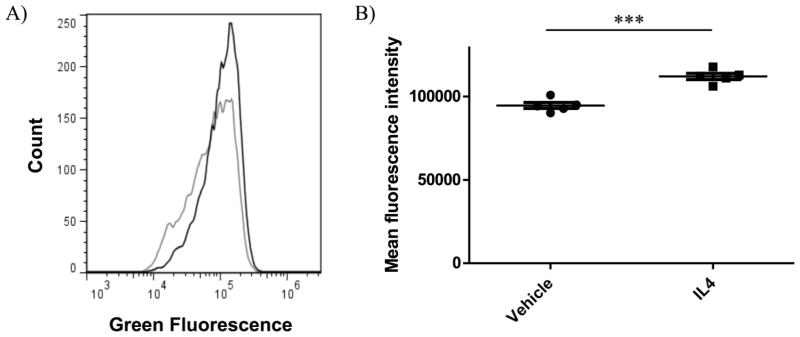
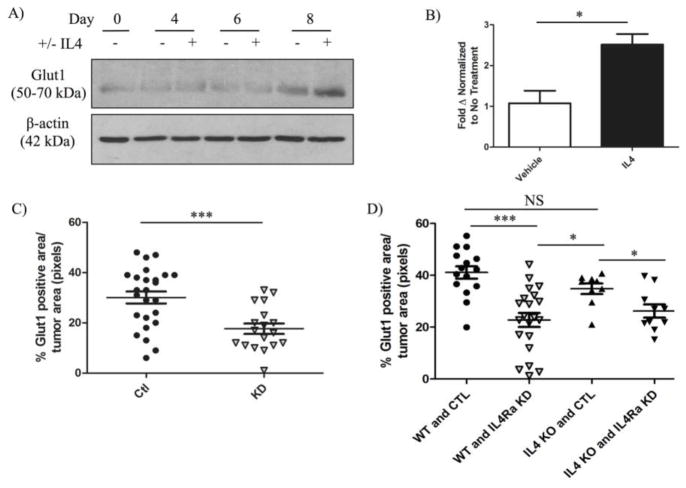
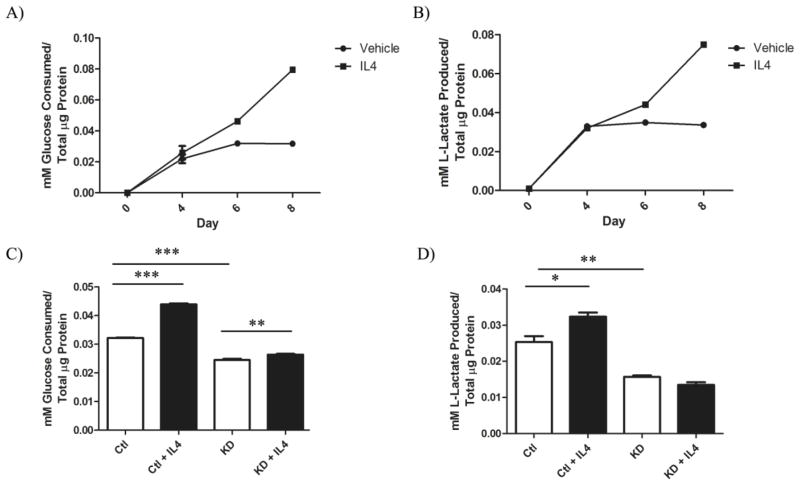
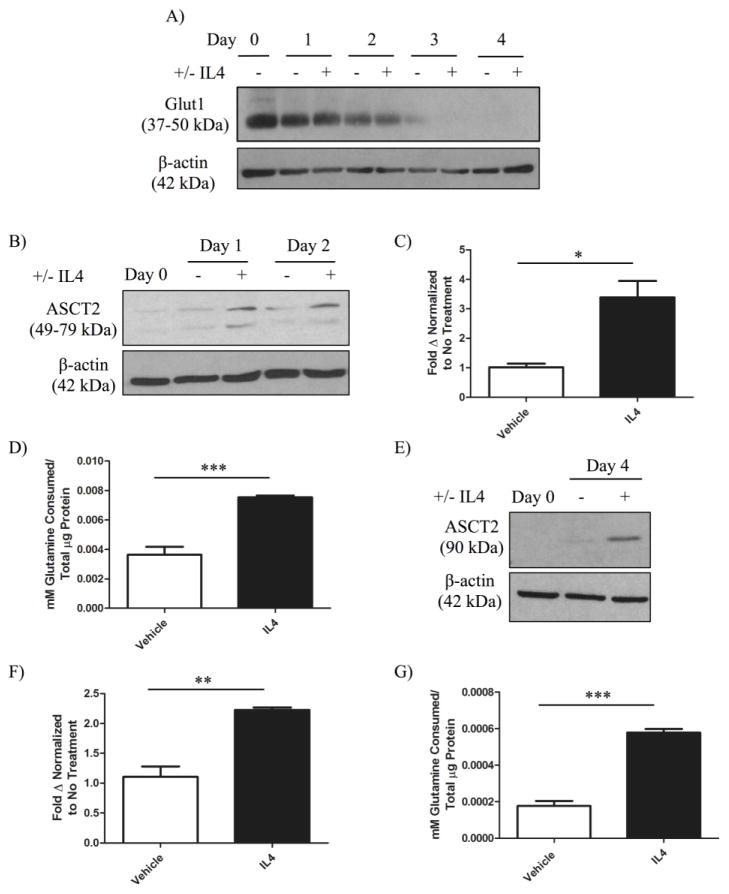
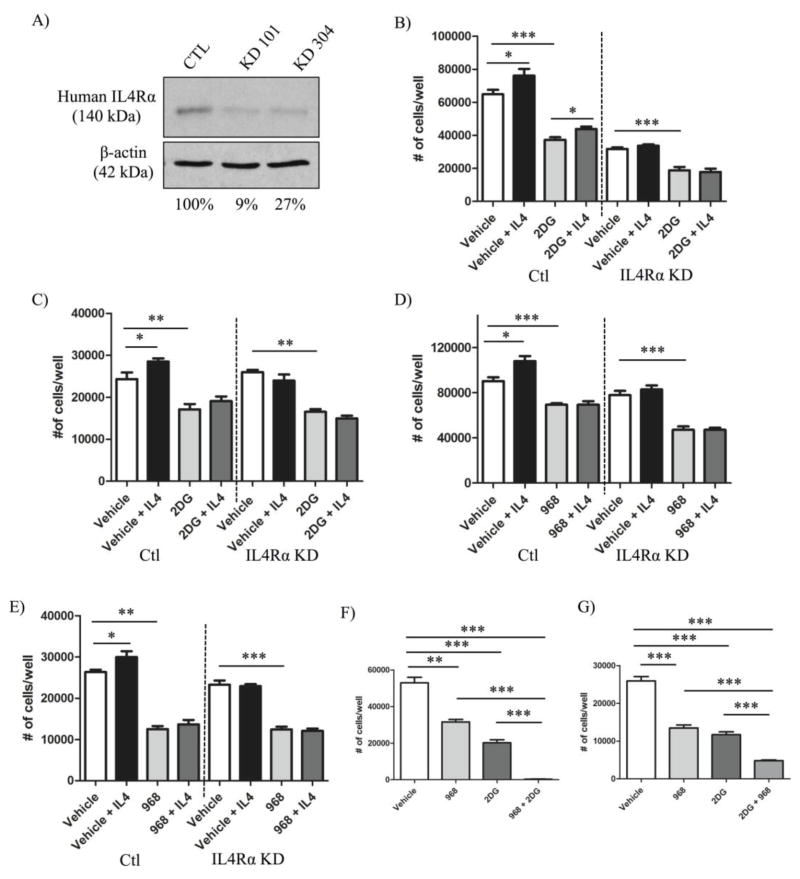
The type II interleukin-4 receptor (IL4R) is expressed in human breast cancer, and in murine models thereof. It is activated by interleukin-4 (IL4), a cytokine produced predominantly by immune cells. Previously, we showed that expression of IL4Rα, a signaling component of IL4R, mediates enhanced metastatic growth through promotion of tumor cell survival and proliferation. In lymphocytes, these processes are supported by increased glucose and glutamine metabolism, and B lymphocyte survival is dependent upon IL4/IL4R-induced glucose metabolism. However, it is unknown whether IL4R-mediated metabolic reprogramming could support tumor growth. Here, we show that IL4Rα expression increases proliferation thus enhancing primary mammary tumor growth. In vitro, IL4-enhanced glucose consumption and lactate production in 4T1 cells was mediated by IL4Rα. Expression of the glucose transporter GLUT1 increased in response to IL4 in vitro, and enhanced GLUT1 expression was associated with the presence of IL4Rα in 4T1 mammary tumors in vivo. Although IL4 treatment did not induce changes in glucose metabolism in MDA-MB-231 human breast cancer cells, it increased expression of the main glutamine transporter, ASCT2, and enhanced glutamine consumption in both MDA-MB-231 and 4T1 cells. Pharmacologic inhibition of glutamine metabolism with compound 968 blocked IL4/IL4Rα-increased cell number in both cell lines. Our results demonstrate that IL4R mediates enhanced glucose and glutamine metabolism in 4T1 cancer cells, and that IL4-induced growth is supported by IL4/IL4R-enhanced glutamine metabolism in both human and murine mammary cancer cells. This highlights IL4Rα as a possible target for effective breast cancer therapy.
Keywords: Cytokine; Glucose; Metabolism; Proliferation; Survival.
Copyright © 2015 Elsevier B.V. All rights reserved.
Conflict of interest statement
No conflicts of interest to declare.
Figures
References
-
- Cancer Facts and Figures 2013. American Cancer Society; Atlanta: 2013.
-
- Nelms K, Keegan AD, Zamorano J, Ryan JJ, Paul WE. The IL-4 receptor: signaling mechanisms and biologic functions. Annu Rev Immunol. 1999;17:701–38. - PubMed
-
- Todaro M, Lombardo Y, Francipane MG, Alea MP, Cammareri P, Iovino F, et al. Apoptosis resistance in epithelial tumors is mediated by tumor-cell-derived interleukin-4. Cell Death Differ. 2008;15:762–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
