

Mutational Analyses of Glucose Dehydrogenase and Glucose-6-Phosphate Dehydrogenase Genes in Pseudomonas fluorescens Reveal Their Effects on Growth and Alginate Production
- PMID: 25746989
- PMCID: PMC4407215
- DOI: 10.1128/AEM.03653-14
Mutational Analyses of Glucose Dehydrogenase and Glucose-6-Phosphate Dehydrogenase Genes in Pseudomonas fluorescens Reveal Their Effects on Growth and Alginate Production
Abstract
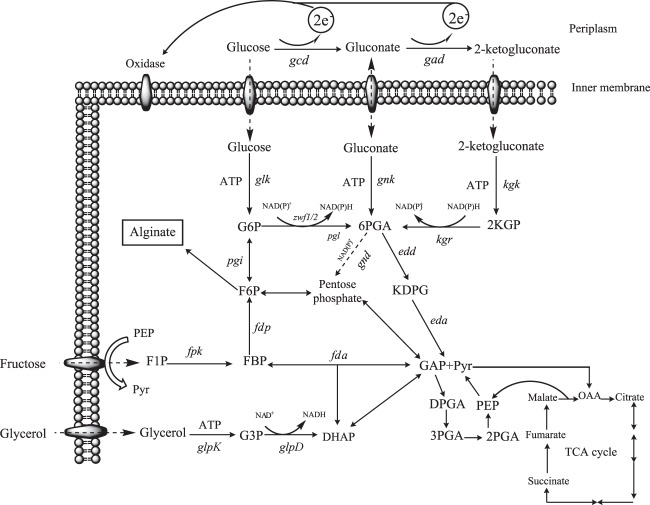
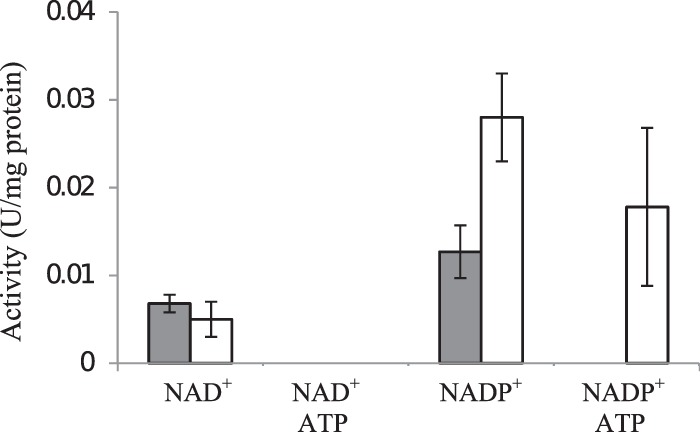
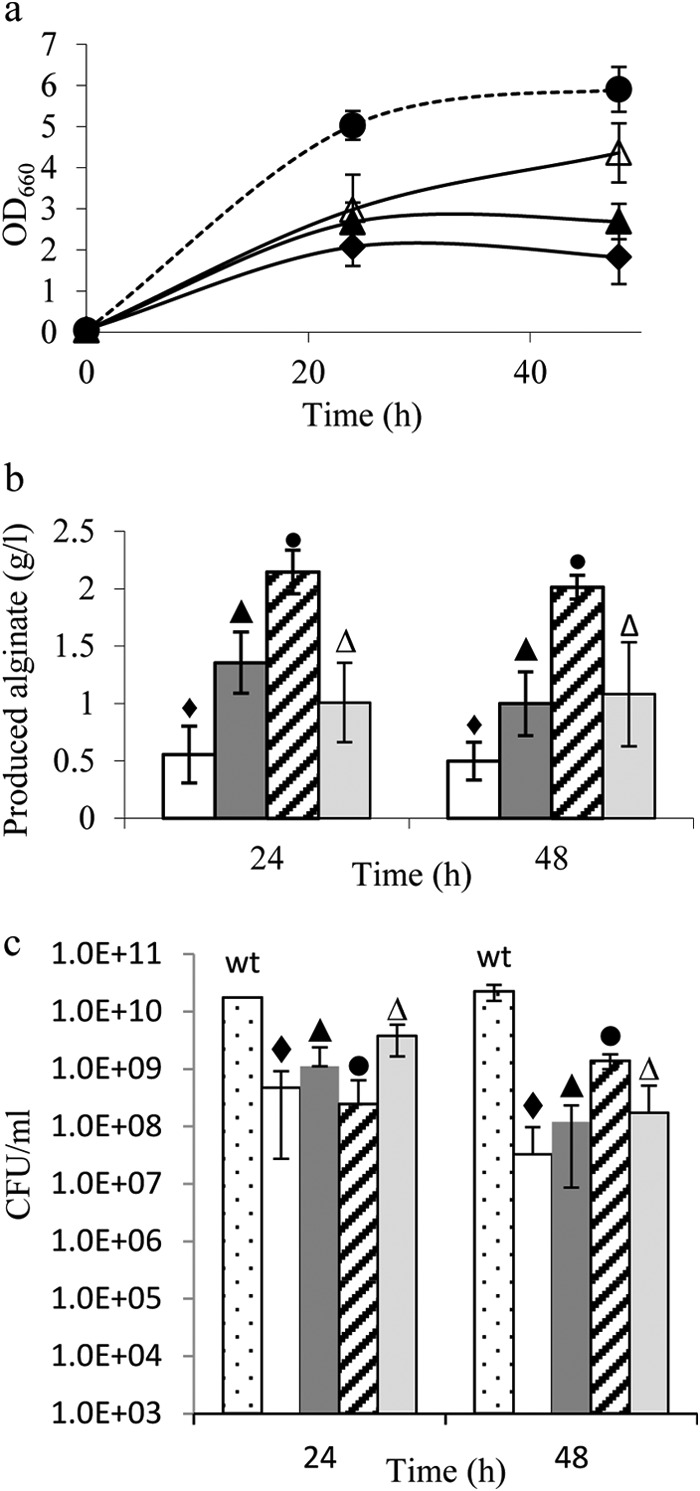
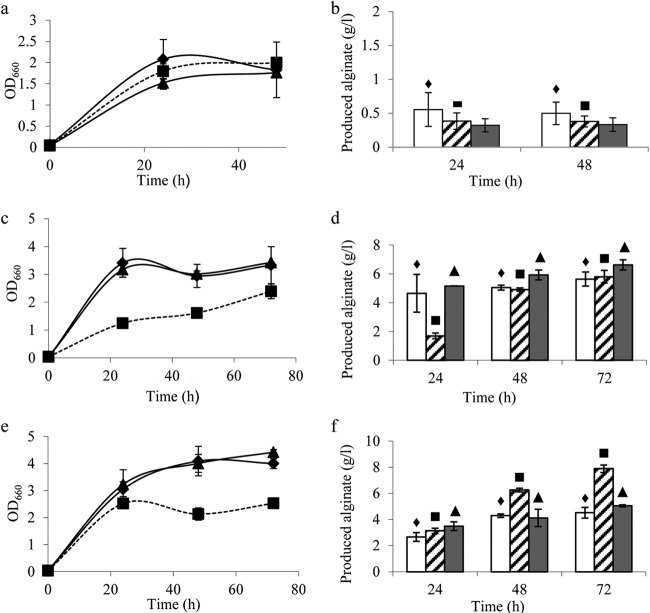
The biosynthesis of alginate has been studied extensively due to the importance of this polymer in medicine and industry. Alginate is synthesized from fructose-6-phosphate and thus competes with the central carbon metabolism for this metabolite. The alginate-producing bacterium Pseudomonas fluorescens relies on the Entner-Doudoroff and pentose phosphate pathways for glucose metabolism, and these pathways are also important for the metabolism of fructose and glycerol. In the present study, the impact of key carbohydrate metabolism enzymes on growth and alginate synthesis was investigated in P. fluorescens. Mutants defective in glucose-6-phosphate dehydrogenase isoenzymes (Zwf-1 and Zwf-2) or glucose dehydrogenase (Gcd) were evaluated using media containing glucose, fructose, or glycerol. Zwf-1 was shown to be the most important glucose-6-phosphate dehydrogenase for catabolism. Both Zwf enzymes preferred NADP as a coenzyme, although NAD was also accepted. Only Zwf-2 was active in the presence of 3 mM ATP, and then only with NADP as a coenzyme, indicating an anabolic role for this isoenzyme. Disruption of zwf-1 resulted in increased alginate production when glycerol was used as the carbon source, possibly due to decreased flux through the Entner-Doudoroff pathway rendering more fructose-6-phosphate available for alginate biosynthesis. In alginate-producing cells grown on glucose, disruption of gcd increased both cell numbers and alginate production levels, while this mutation had no positive effect on growth in a non-alginate-producing strain. A possible explanation is that alginate synthesis might function as a sink for surplus hexose phosphates that could otherwise be detrimental to the cell.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures

Similar articles
-
New insights into Pseudomonas fluorescens alginate biosynthesis relevant for the establishment of an efficient production process for microbial alginates.N Biotechnol. 2017 Jul 25;37(Pt A):2-8. doi: 10.1016/j.nbt.2016.08.005. Epub 2016 Sep 1. N Biotechnol. 2017. PMID: 27593394 Review.
-
Identification of a New Phosphatase Enzyme Potentially Involved in the Sugar Phosphate Stress Response in Pseudomonas fluorescens.Appl Environ Microbiol. 2016 Dec 30;83(2):e02361-16. doi: 10.1128/AEM.02361-16. Print 2017 Jan 15. Appl Environ Microbiol. 2016. PMID: 27836849 Free PMC article.
-
Identification of genes affecting alginate biosynthesis in Pseudomonas fluorescens by screening a transposon insertion library.BMC Genomics. 2017 Jan 3;18(1):11. doi: 10.1186/s12864-016-3467-7. BMC Genomics. 2017. PMID: 28049432 Free PMC article.
-
Adaptations of Pseudomonas aeruginosa to the cystic fibrosis lung environment can include deregulation of zwf, encoding glucose-6-phosphate dehydrogenase.J Bacteriol. 2005 Nov;187(22):7561-8. doi: 10.1128/JB.187.22.7561-7568.2005. J Bacteriol. 2005. PMID: 16267280 Free PMC article.
-
Glucose-6-phosphate dehydrogenase is required for extracellular polysaccharide production, cell motility and the full virulence of Xanthomonas oryzae pv. oryzicola.Microb Pathog. 2015 Jan;78:87-94. doi: 10.1016/j.micpath.2014.11.007. Epub 2014 Nov 18. Microb Pathog. 2015. PMID: 25450881
Cited by
-
Pseudomonas aeruginosa Utilizes Host-Derived Itaconate to Redirect Its Metabolism to Promote Biofilm Formation.Cell Metab. 2020 Jun 2;31(6):1091-1106.e6. doi: 10.1016/j.cmet.2020.04.017. Epub 2020 May 18. Cell Metab. 2020. PMID: 32428444 Free PMC article.
-
Identification of Regulatory Genes and Metabolic Processes Important for Alginate Biosynthesis in Azotobacter vinelandii by Screening of a Transposon Insertion Mutant Library.Front Bioeng Biotechnol. 2020 Jan 17;7:475. doi: 10.3389/fbioe.2019.00475. eCollection 2019. Front Bioeng Biotechnol. 2020. PMID: 32010681 Free PMC article.
-
Fused Enzyme Glucose-6-Phosphate Dehydrogenase::6-Phosphogluconolactonase (G6PD::6PGL) as a Potential Drug Target in Giardia lamblia, Trichomonas vaginalis, and Plasmodium falciparum.Microorganisms. 2024 Jan 5;12(1):112. doi: 10.3390/microorganisms12010112. Microorganisms. 2024. PMID: 38257939 Free PMC article. Review.
-
Pseudomonas putida KT2440 Strain Metabolizes Glucose through a Cycle Formed by Enzymes of the Entner-Doudoroff, Embden-Meyerhof-Parnas, and Pentose Phosphate Pathways.J Biol Chem. 2015 Oct 23;290(43):25920-32. doi: 10.1074/jbc.M115.687749. Epub 2015 Sep 8. J Biol Chem. 2015. PMID: 26350459 Free PMC article.
-
Alginate Biosynthesis Factories in Pseudomonas fluorescens: Localization and Correlation with Alginate Production Level.Appl Environ Microbiol. 2015 Dec 11;82(4):1227-1236. doi: 10.1128/AEM.03114-15. Print 2016 Feb 15. Appl Environ Microbiol. 2015. PMID: 26655760 Free PMC article.
References
-
- Gimmestad M, Sletta H, Karunakaran P, Bakkevig K, Ertesvåg H, Ellingsen TE, Skjåk-Bræk G, Valla S. 7 May 2014. New mutant strains of Pseudomonas fluorescens and variants thereof, methods of their production, and uses thereof in alginate production. European patent WO 2004/011628.
-
- Lessmann D, Schimz KL, Kurz G. 1975. d-Glucose-6-phosphate dehydrogenase (Entner-Doudoroff enzyme) from Pseudomonas fluorescens. Purification, properties and regulation. Eur J Biochem 59:545–559. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
