

Comparing different stimulus configurations for population receptive field mapping in human fMRI
- PMID: 25750620
- PMCID: PMC4335485
- DOI: 10.3389/fnhum.2015.00096
Comparing different stimulus configurations for population receptive field mapping in human fMRI
Abstract
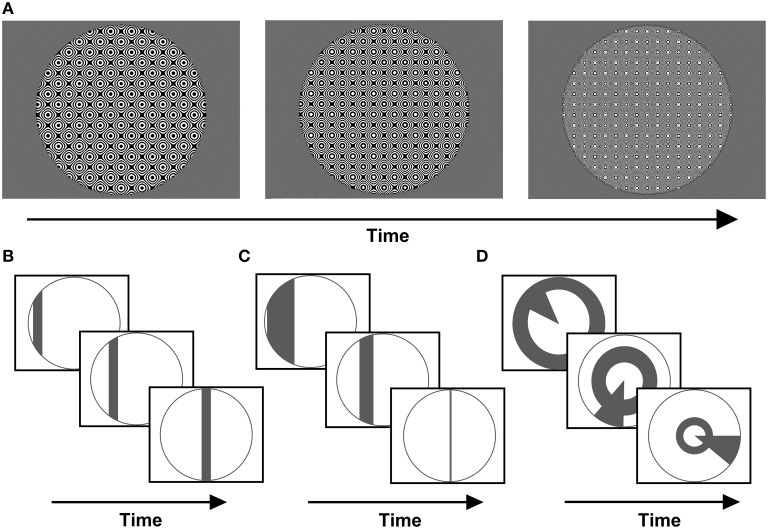
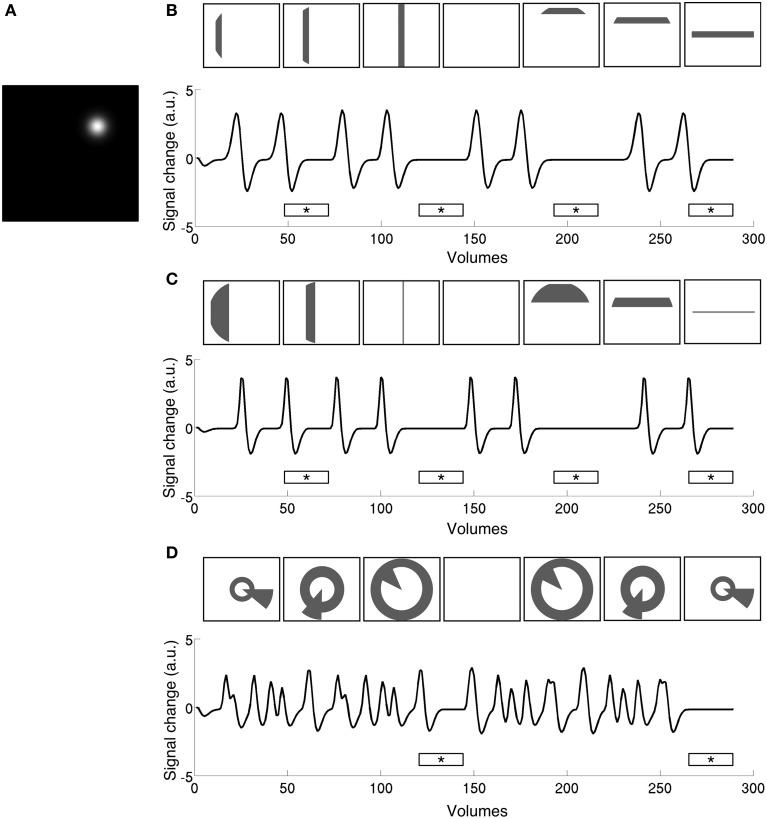
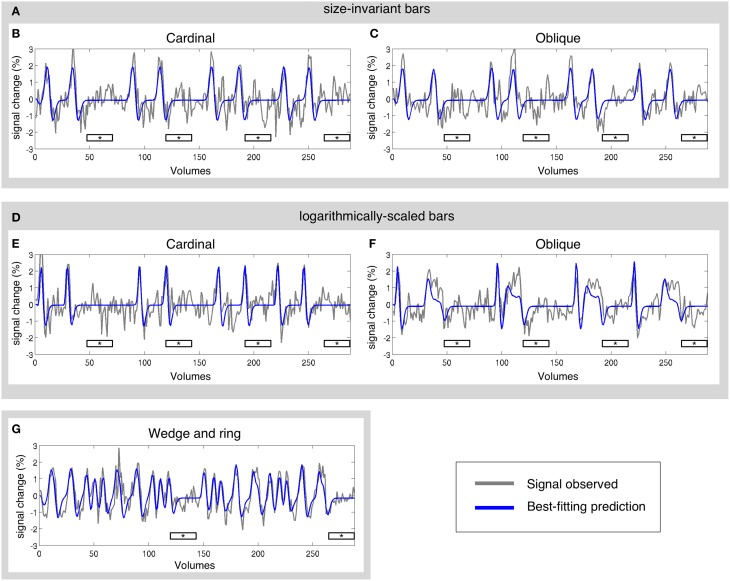
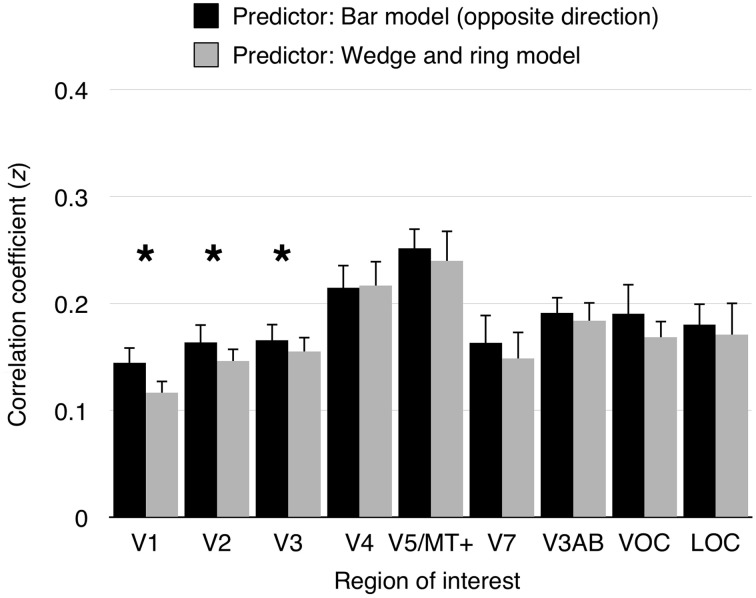
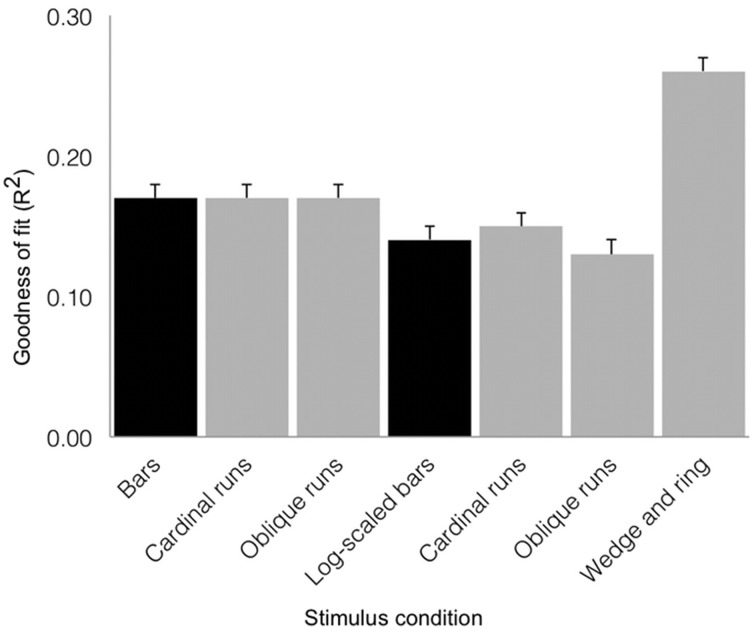
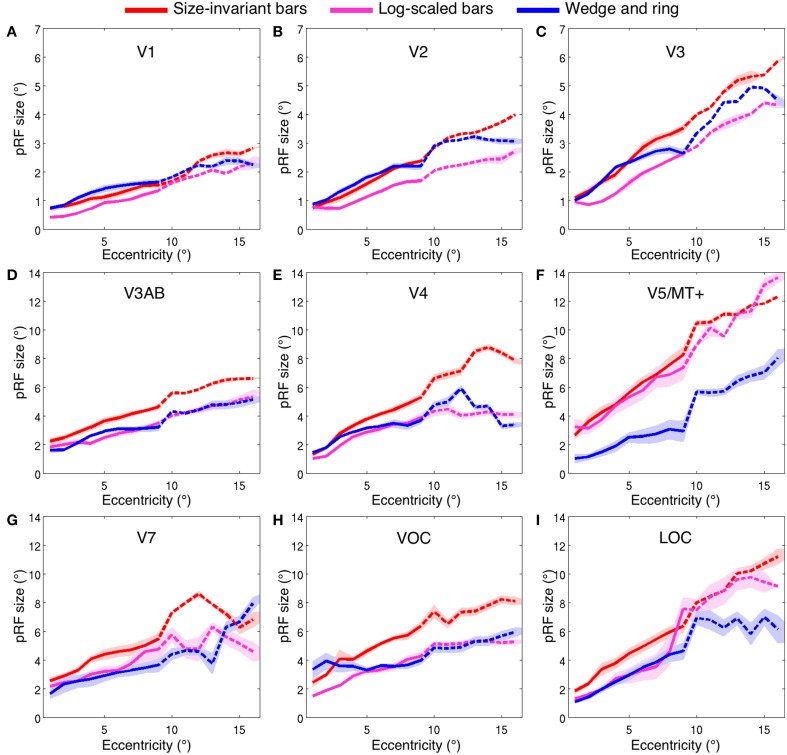
Population receptive field (pRF) mapping is a widely used approach to measuring aggregate human visual receptive field properties by recording non-invasive signals using functional MRI. Despite growing interest, no study to date has systematically investigated the effects of different stimulus configurations on pRF estimates from human visual cortex. Here we compared the effects of three different stimulus configurations on a model-based approach to pRF estimation: size-invariant bars and eccentricity-scaled bars defined in Cartesian coordinates and traveling along the cardinal axes, and a novel simultaneous "wedge and ring" stimulus defined in polar coordinates, systematically covering polar and eccentricity axes. We found that the presence or absence of eccentricity scaling had a significant effect on goodness of fit and pRF size estimates. Further, variability in pRF size estimates was directly influenced by stimulus configuration, particularly for higher visual areas including V5/MT+. Finally, we compared eccentricity estimation between phase-encoded and model-based pRF approaches. We observed a tendency for more peripheral eccentricity estimates using phase-encoded methods, independent of stimulus size. We conclude that both eccentricity scaling and polar rather than Cartesian stimulus configuration are important considerations for optimal experimental design in pRF mapping. While all stimulus configurations produce adequate estimates, simultaneous wedge and ring stimulation produced higher fit reliability, with a significant advantage in reduced acquisition time.
Keywords: fMRI; pRF; population receptive field modeling; primary visual cortex (V1); retinotopy; stimulus design; visual cortex.
Figures


References
-
- Ashburner J., Friston K. J. (1997). Spatial transformation of images, in Human Brain Function, eds Frackowiak R. S., Friston K. J., Frith C. D., Dolan R. J., Mazziotta J. C. (Waltham, MA: Academic Press; ), 43–58.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources