

Evidence that processing of ribonucleotides in DNA by topoisomerase 1 is leading-strand specific
- PMID: 25751426
- PMCID: PMC4835660
- DOI: 10.1038/nsmb.2989
Evidence that processing of ribonucleotides in DNA by topoisomerase 1 is leading-strand specific
Abstract
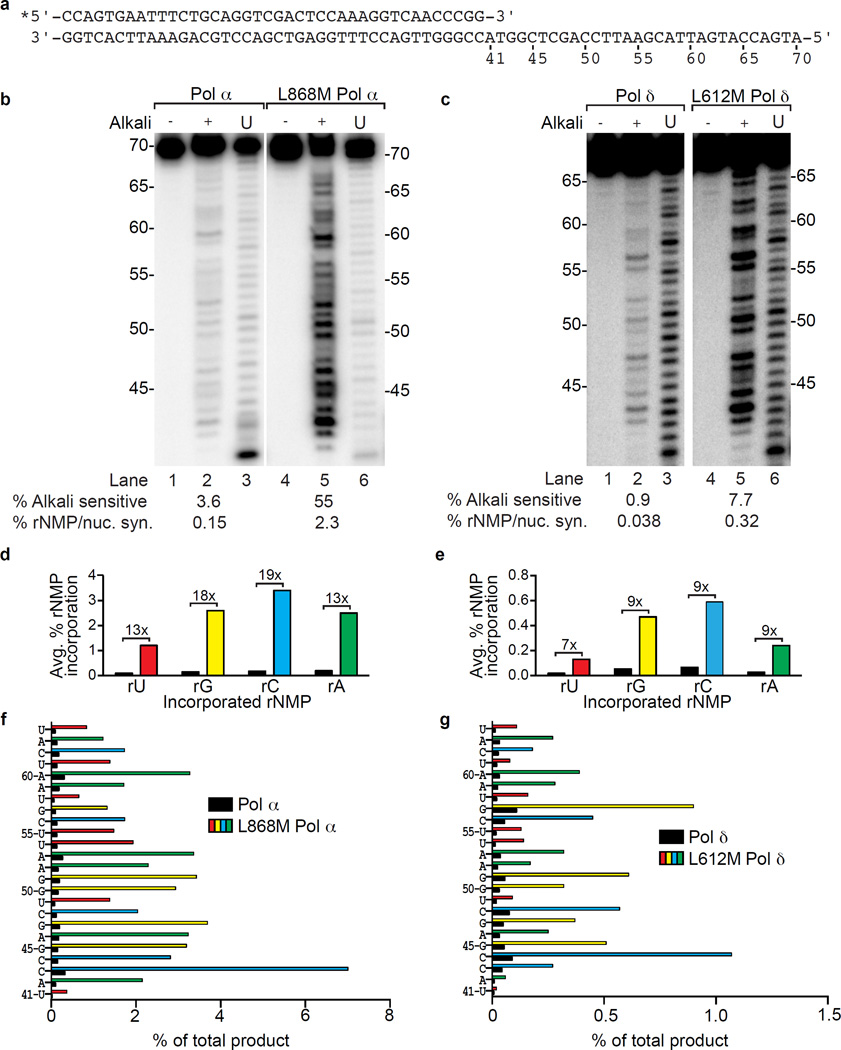
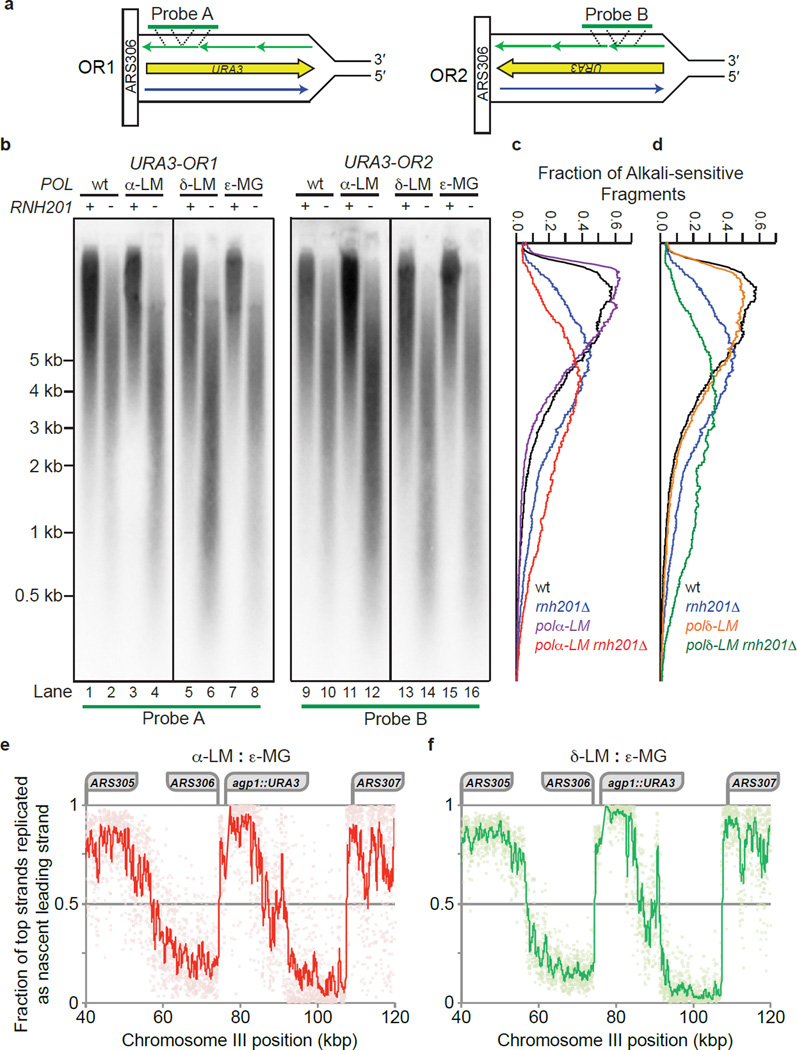
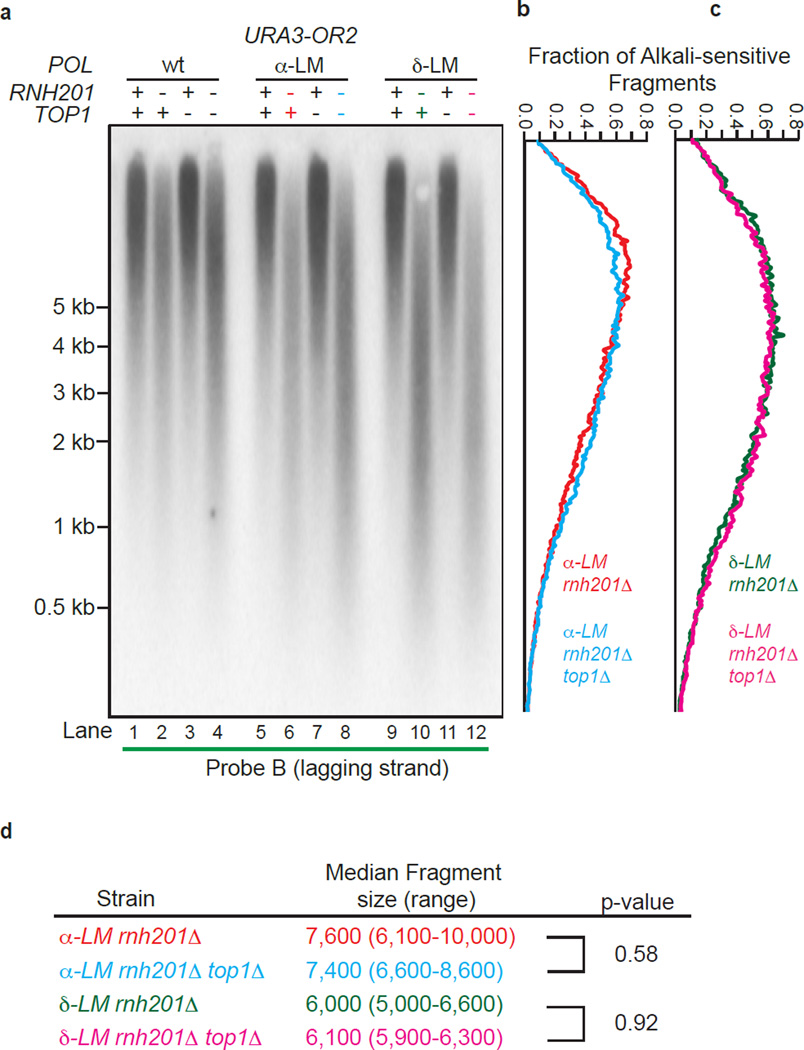
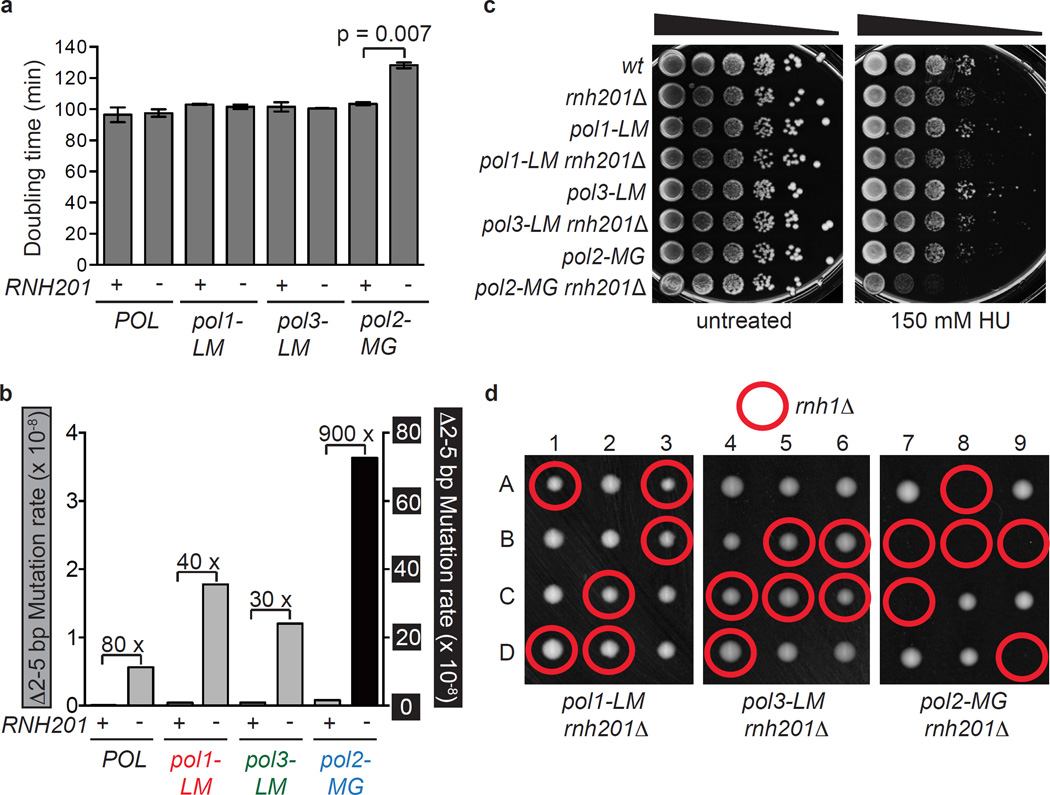
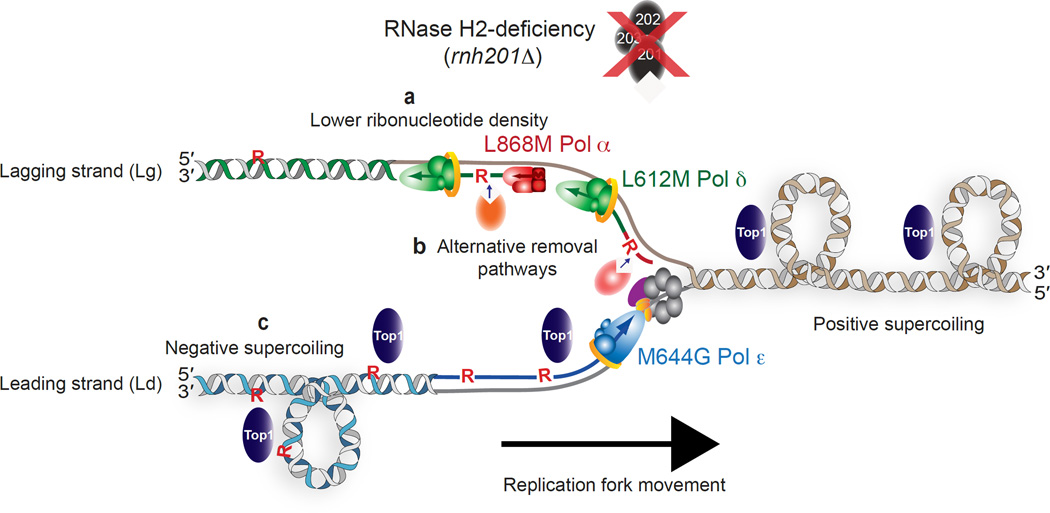
Ribonucleotides incorporated during DNA replication are removed by RNase H2-dependent ribonucleotide excision repair (RER). In RER-defective yeast, topoisomerase 1 (Top1) incises DNA at unrepaired ribonucleotides, initiating their removal, but this is accompanied by RNA-DNA-damage phenotypes. Here we show that these phenotypes are incurred by a high level of ribonucleotides incorporated by a leading strand-replicase variant, DNA polymerase (Pol) ɛ, but not by orthologous variants of the lagging-strand replicases, Pols α or δ. Moreover, loss of both RNases H1 and H2 is lethal in combination with increased ribonucleotide incorporation by Pol ɛ but not by Pols α or δ. Several explanations for this asymmetry are considered, including the idea that Top1 incision at ribonucleotides relieves torsional stress in the nascent leading strand but not in the nascent lagging strand, in which preexisting nicks prevent the accumulation of superhelical tension.
Figures
References
-
- Wang J, et al. Crystal structure of a pol alpha family replication DNA polymerase from bacteriophage RB69. Cell. 1997;89:1087–1099. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
