

Variable Substrate Preference among Phospholipase D Toxins from Sicariid Spiders
- PMID: 25752604
- PMCID: PMC4409260
- DOI: 10.1074/jbc.M115.636951
Variable Substrate Preference among Phospholipase D Toxins from Sicariid Spiders
Abstract
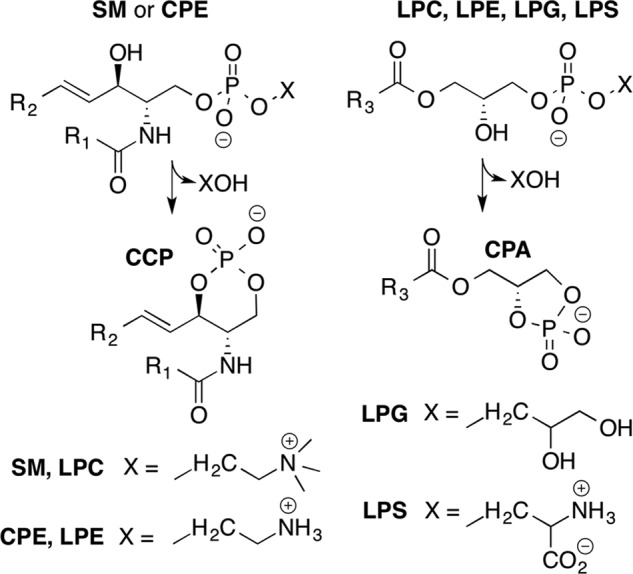
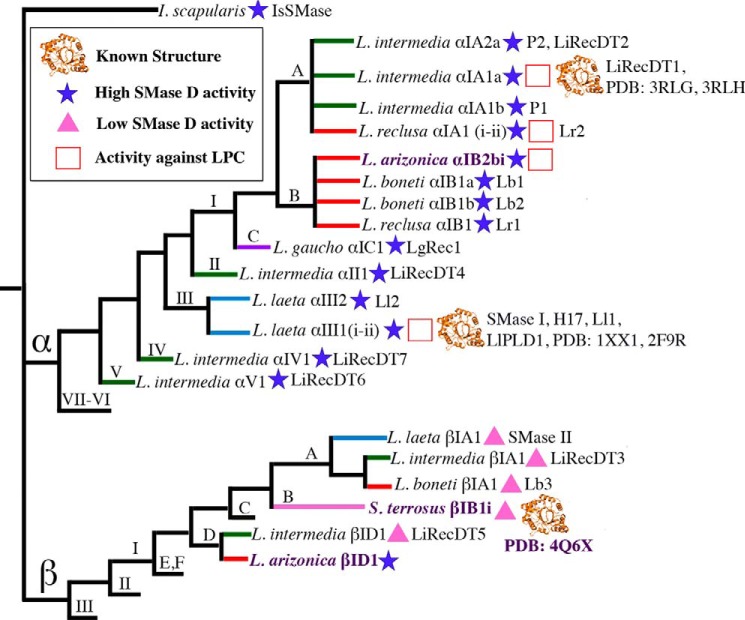
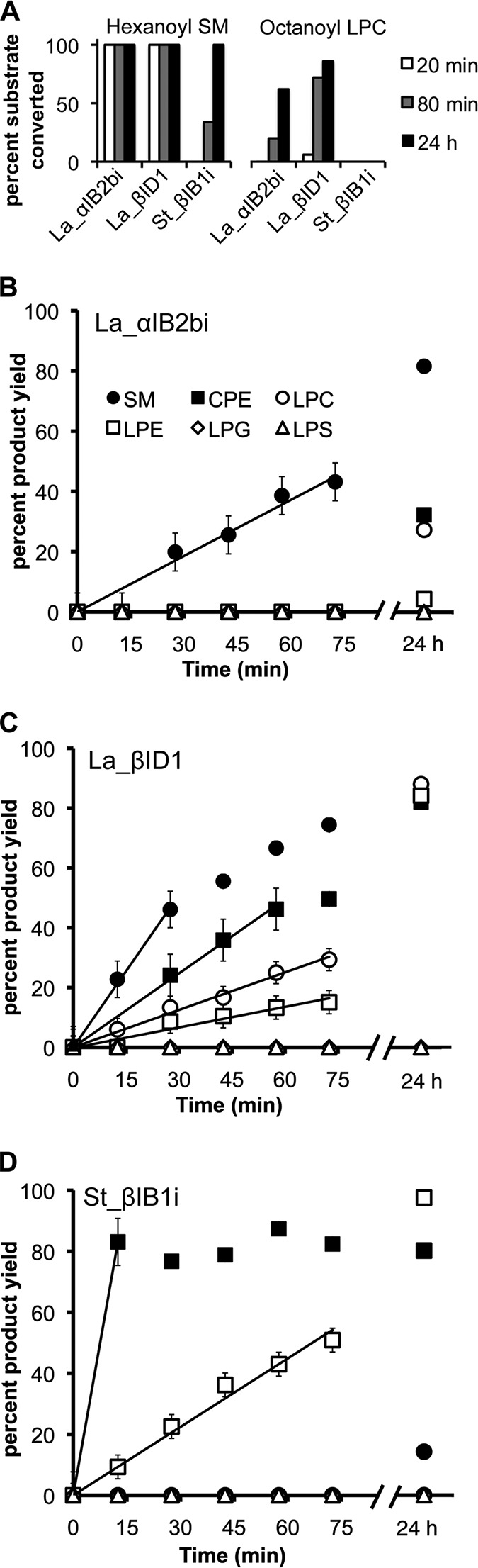
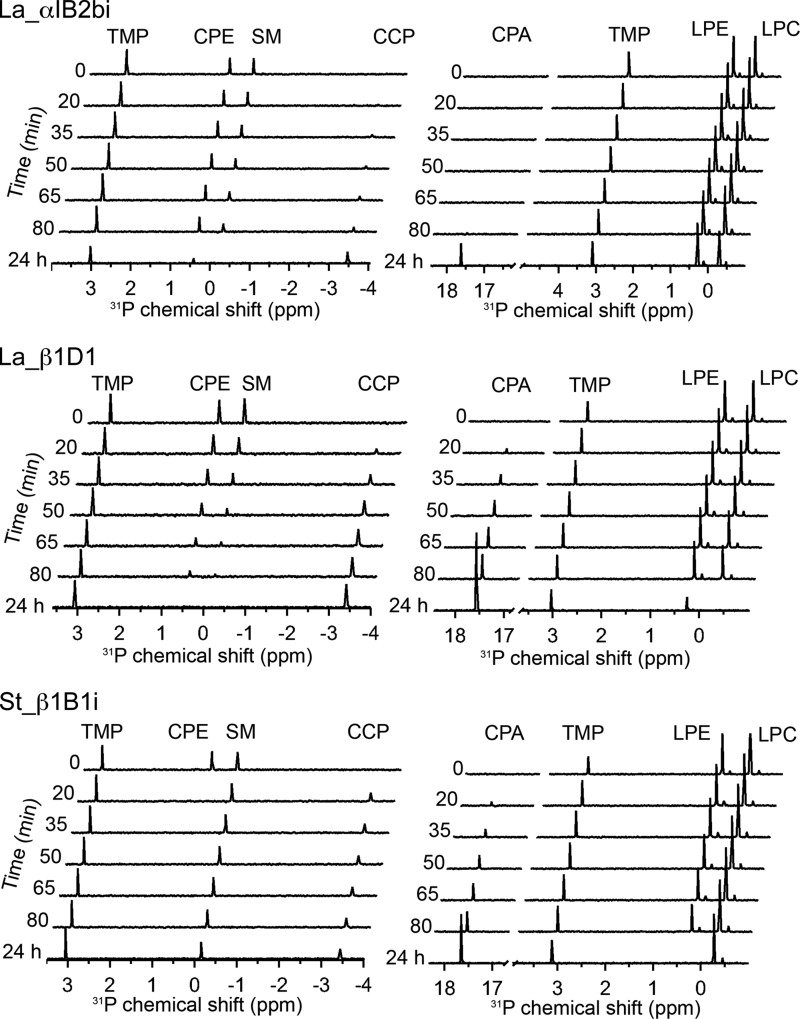
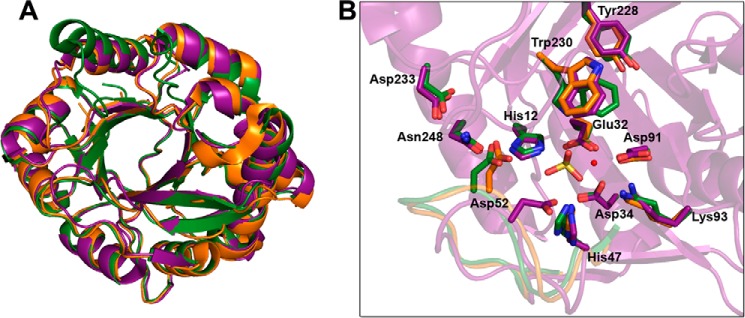
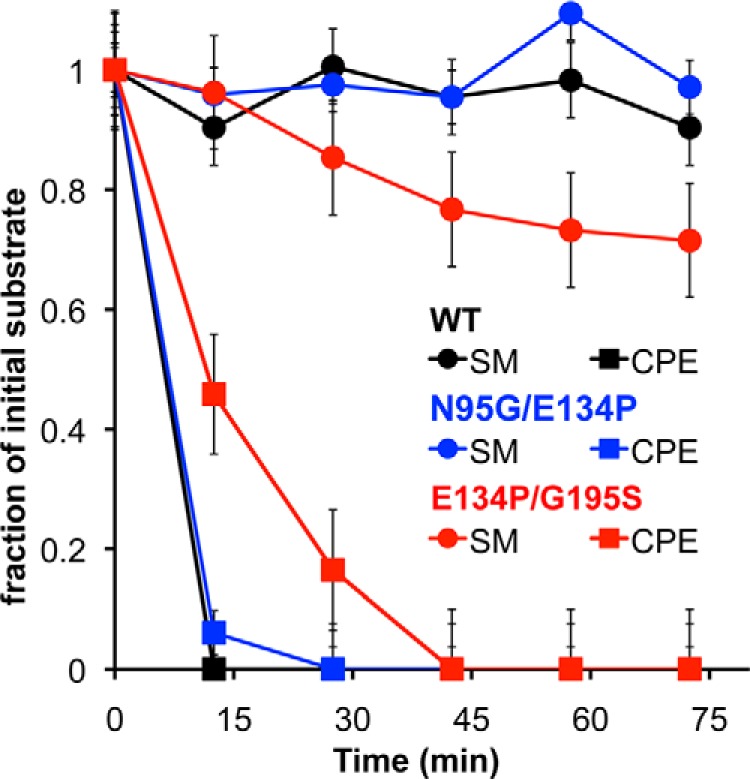
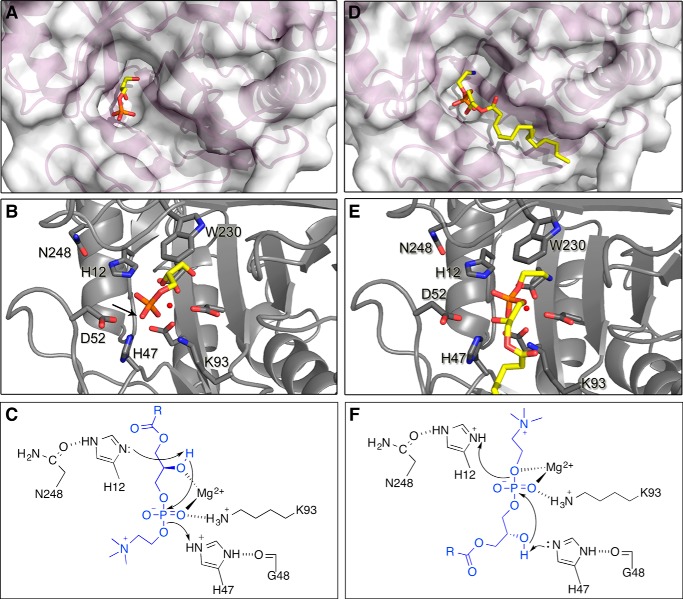
Venoms of the sicariid spiders contain phospholipase D enzyme toxins that can cause severe dermonecrosis and even death in humans. These enzymes convert sphingolipid and lysolipid substrates to cyclic phosphates by activating a hydroxyl nucleophile present in both classes of lipid. The most medically relevant substrates are thought to be sphingomyelin and/or lysophosphatidylcholine. To better understand the substrate preference of these toxins, we used (31)P NMR to compare the activity of three related but phylogenetically diverse sicariid toxins against a diverse panel of sphingolipid and lysolipid substrates. Two of the three showed significantly faster turnover of sphingolipids over lysolipids, and all three showed a strong preference for positively charged (choline and/or ethanolamine) over neutral (glycerol and serine) headgroups. Strikingly, however, the enzymes vary widely in their preference for choline, the headgroup of both sphingomyelin and lysophosphatidylcholine, versus ethanolamine. An enzyme from Sicarius terrosus showed a strong preference for ethanolamine over choline, whereas two paralogous enzymes from Loxosceles arizonica either preferred choline or showed no significant preference. Intrigued by the novel substrate preference of the Sicarius enzyme, we solved its crystal structure at 2.1 Å resolution. The evolution of variable substrate specificity may help explain the reduced dermonecrotic potential of some natural toxin variants, because mammalian sphingolipids use primarily choline as a positively charged headgroup; it may also be relevant for sicariid predatory behavior, because ethanolamine-containing sphingolipids are common in insect prey.
Keywords: Crystal Structure; Enzyme Mechanism; Loxosceles; Loxoscelism; Phospholipase D; Sicarius; Sphingomyelinase; Substrate Specificity; Toxin.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures


References
-
- Tambourgi D. V., Gonçalves-de-Andrade R. M., van den Berg C. W. (2010) Loxoscelism: From basic research to the proposal of new therapies. Toxicon 56, 1113–1119 - PubMed
-
- Gremski L. H., Trevisan-Silva D., Ferrer V. P., Matsubara F. H., Meissner G. O., Wille A. C., Vuitika L., Dias-Lopes C., Ullah A., de Moraes F. R., Chávez-Olortegui C., Barbaro K. C., Murakami M. T., Arni R. K., Senff-Ribeiro A., et al. (2014) Recent advances in the understanding of brown spider venoms: from the biology of spiders to the molecular mechanisms of toxins. Toxicon 83, 91–120 - PubMed
-
- Hogan C. J., Barbaro K. C., Winkel K. (2004) Loxoscelism: old obstacles, new directions. Ann. Emerg. Med. 44, 608–624 - PubMed
-
- da Silva P. H., da Silveira R. B., Appel M. H., Mangili O. C., Gremski W., Veiga S. S. (2004) Brown spiders and loxoscelism. Toxicon 44, 693–709 - PubMed
-
- Swanson D. L., Vetter R. S. (2006) Loxoscelism. Clin. Dermatol. 24, 213–221 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
