

GGCX and VKORC1 inhibit osteocalcin endocrine functions
- PMID: 25753038
- PMCID: PMC4362468
- DOI: 10.1083/jcb.201409111
GGCX and VKORC1 inhibit osteocalcin endocrine functions
Erratum in
-
Correction: GGCX and VKORC1 inhibit osteocalcin endocrine functions.J Cell Biol. 2019 Jun 3;218(6):2071. doi: 10.1083/jcb.20140911104082019c. Epub 2019 Apr 30. J Cell Biol. 2019. PMID: 31040164 Free PMC article. No abstract available.
Abstract
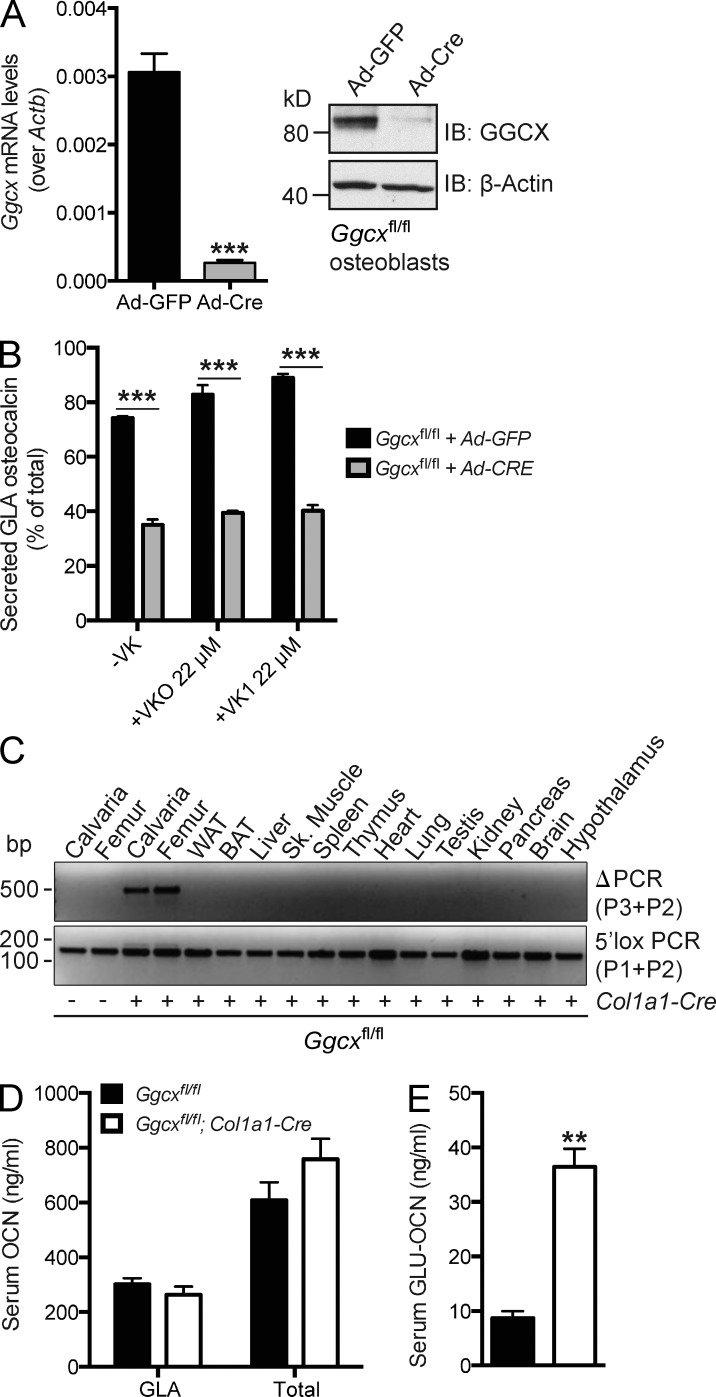
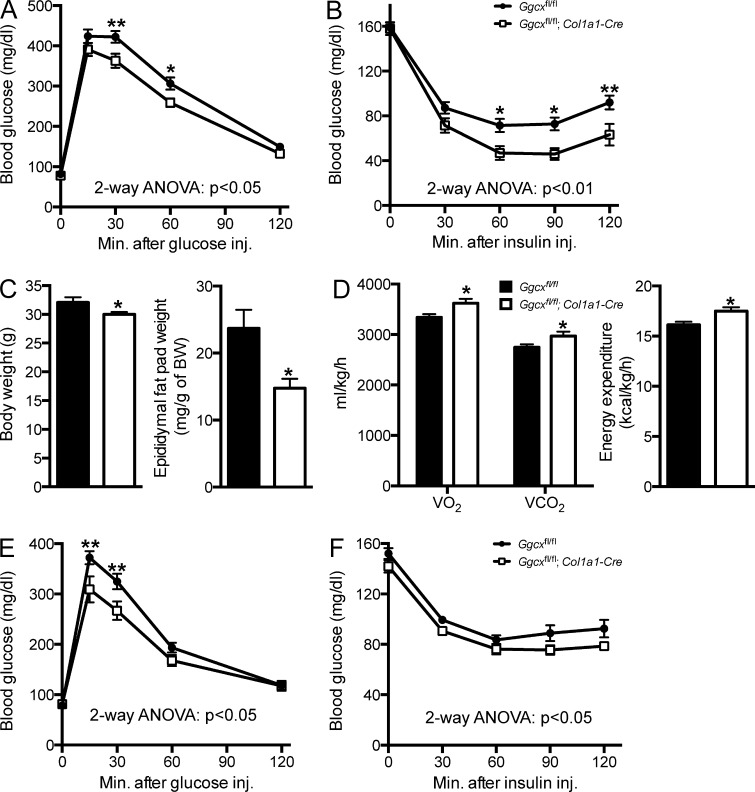
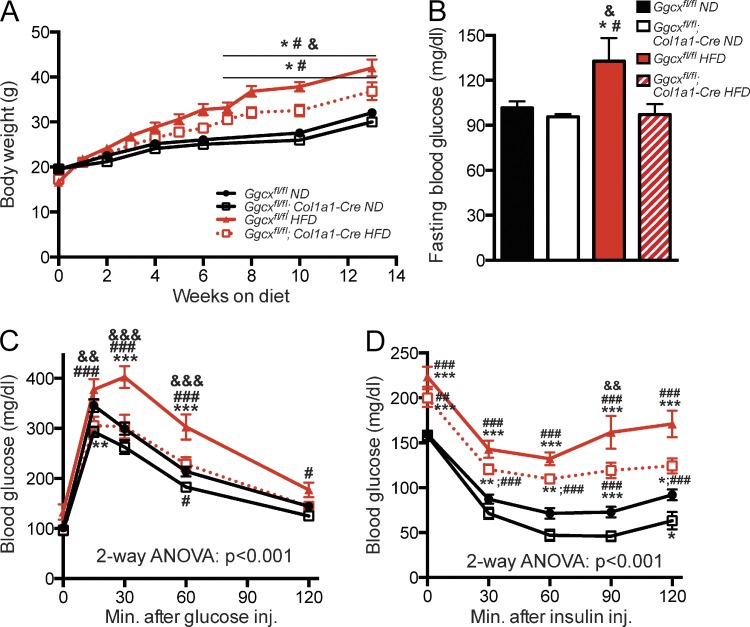
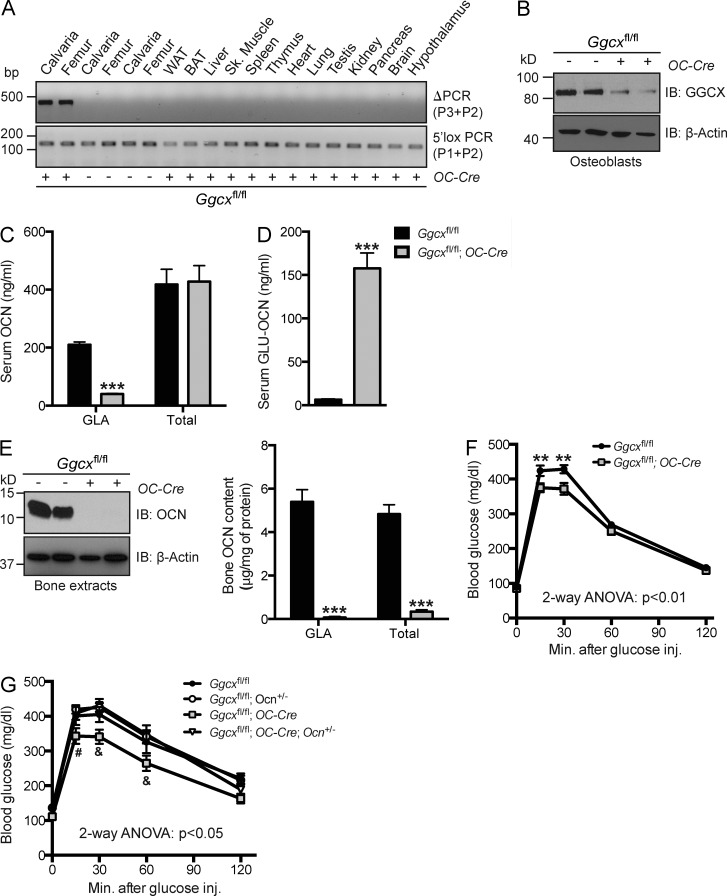
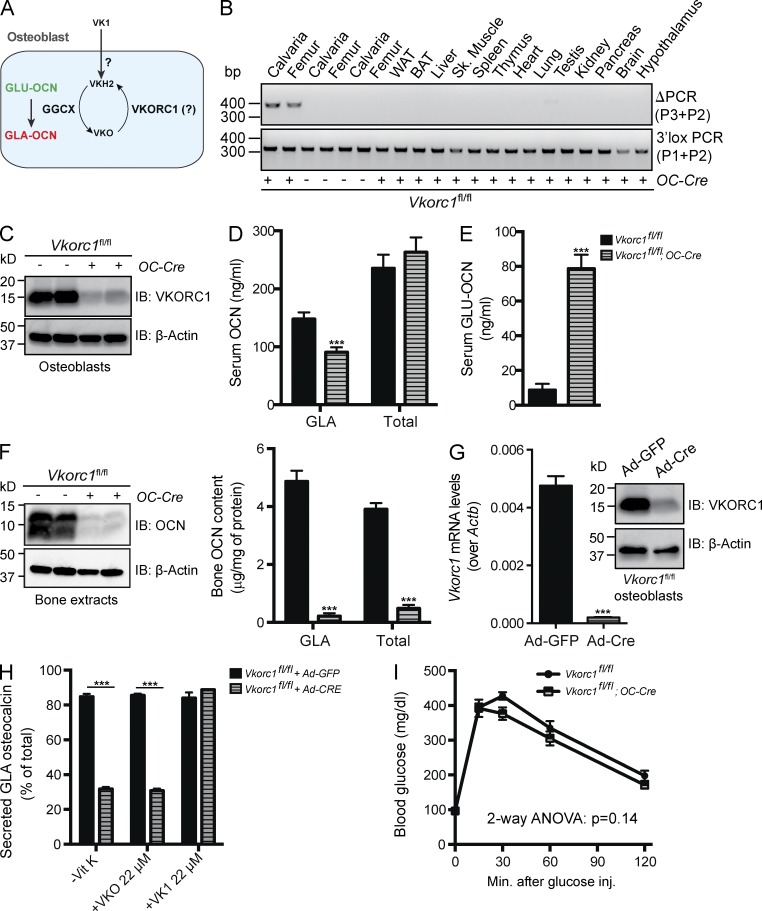
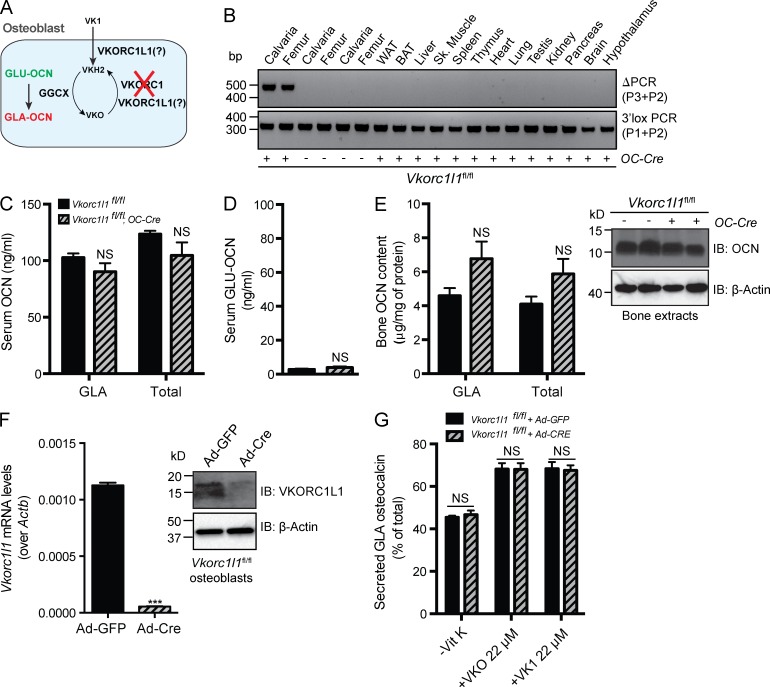
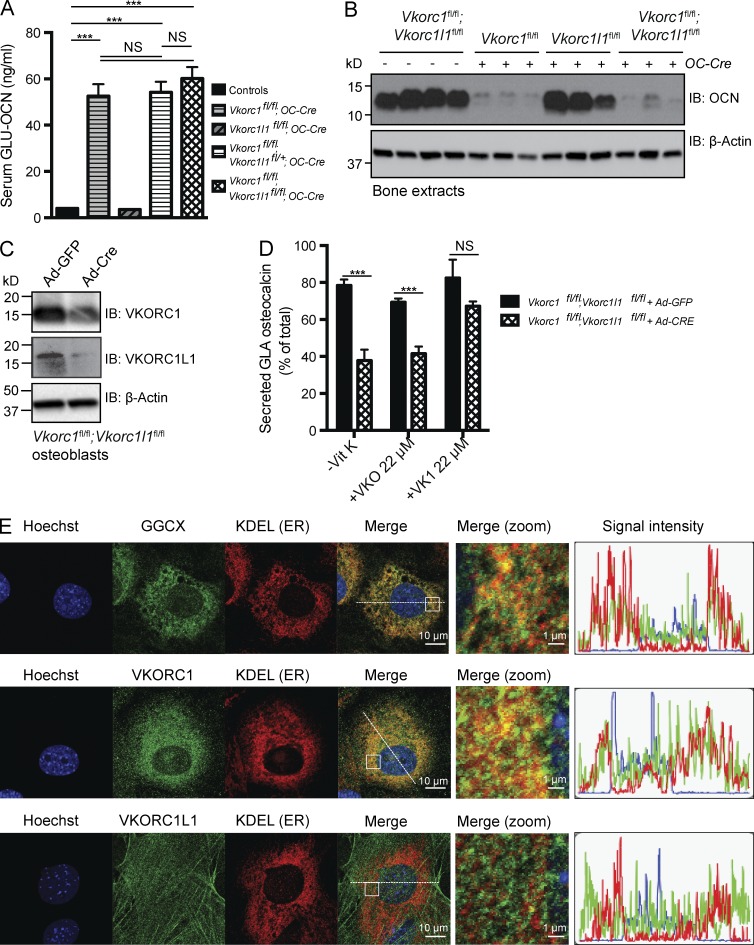
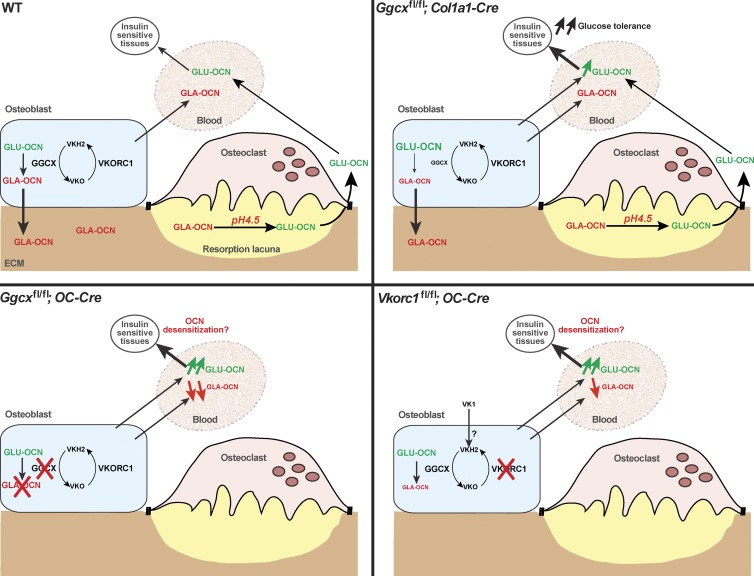
Osteocalcin (OCN) is an osteoblast-derived hormone favoring glucose homeostasis, energy expenditure, male fertility, brain development, and cognition. Before being secreted by osteoblasts in the bone extracellular matrix, OCN is γ-carboxylated by the γ-carboxylase (GGCX) on three glutamic acid residues, a cellular process requiring reduction of vitamin K (VK) by a second enzyme, a reductase called VKORC1. Although circumstantial evidence suggests that γ-carboxylation may inhibit OCN endocrine functions, genetic evidence that it is the case is still lacking. Here we show using cell-specific gene inactivation models that γ-carboxylation of OCN by GGCX inhibits its endocrine function. We further show that VKORC1 is required for OCN γ-carboxylation in osteoblasts, whereas its paralogue, VKORC1L1, is dispensable for this function and cannot compensate for the absence of VKORC1 in osteoblasts. This study genetically and biochemically delineates the functions of the enzymes required for OCN modification and demonstrates that it is the uncarboxylated form of OCN that acts as a hormone.
© 2015 Ferron et al.
Figures
References
-
- Abseyi N., Şıklar Z., Berberoğlu M., Hacıhamdioğlu B., Savaş Erdeve Ş., and Öçal G.. 2012. Relationships between osteocalcin, glucose metabolism, and adiponectin in obese children: Is there crosstalk between bone tissue and glucose metabolism? J. Clin. Res. Pediatr. Endocrinol. 4:182–188. 10.4274/Jcrpe.831 - DOI - PMC - PubMed
-
- Brennan-Speranza T.C., Henneicke H., Gasparini S.J., Blankenstein K.I., Heinevetter U., Cogger V.C., Svistounov D., Zhang Y., Cooney G.J., Buttgereit F., et al. 2012. Osteoblasts mediate the adverse effects of glucocorticoids on fuel metabolism. J. Clin. Invest. 122:4172–4189. 10.1172/JCI63377 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
