

ZO-1 controls endothelial adherens junctions, cell-cell tension, angiogenesis, and barrier formation
- PMID: 25753039
- PMCID: PMC4362456
- DOI: 10.1083/jcb.201404140
ZO-1 controls endothelial adherens junctions, cell-cell tension, angiogenesis, and barrier formation
Abstract
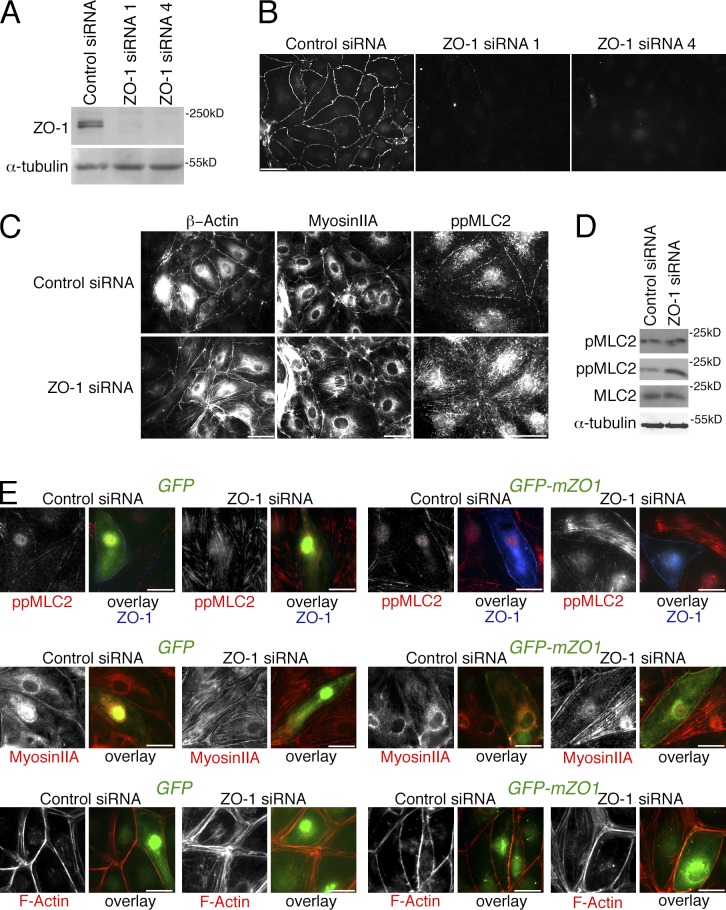
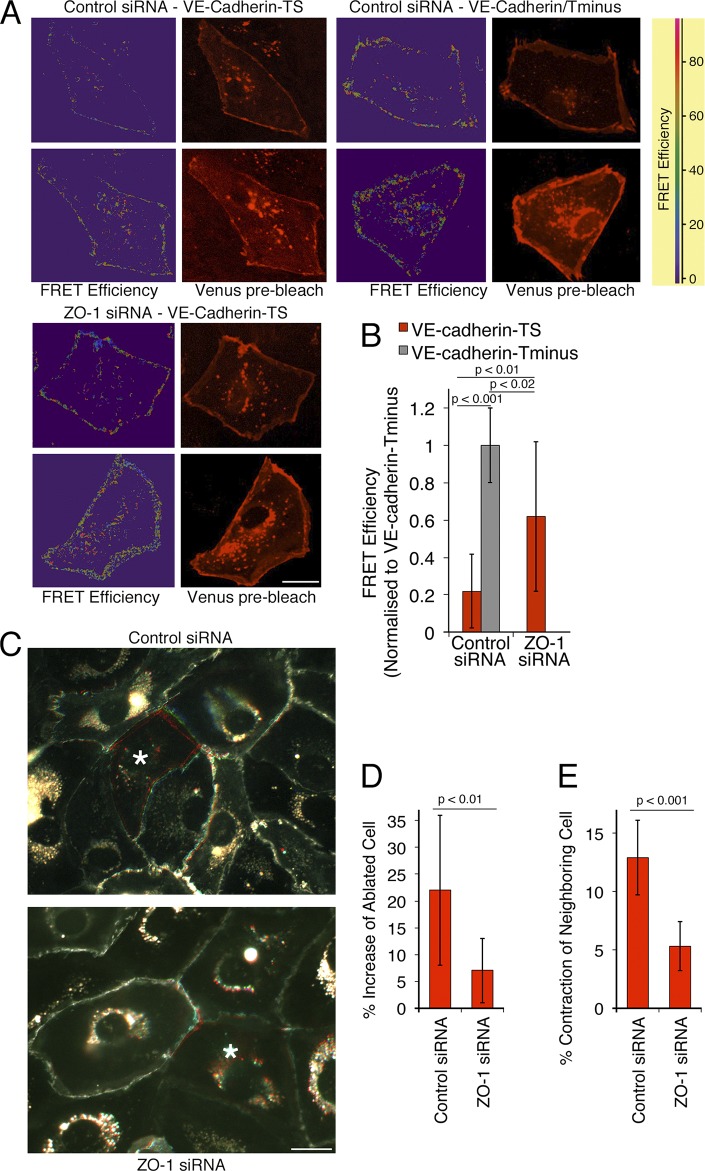
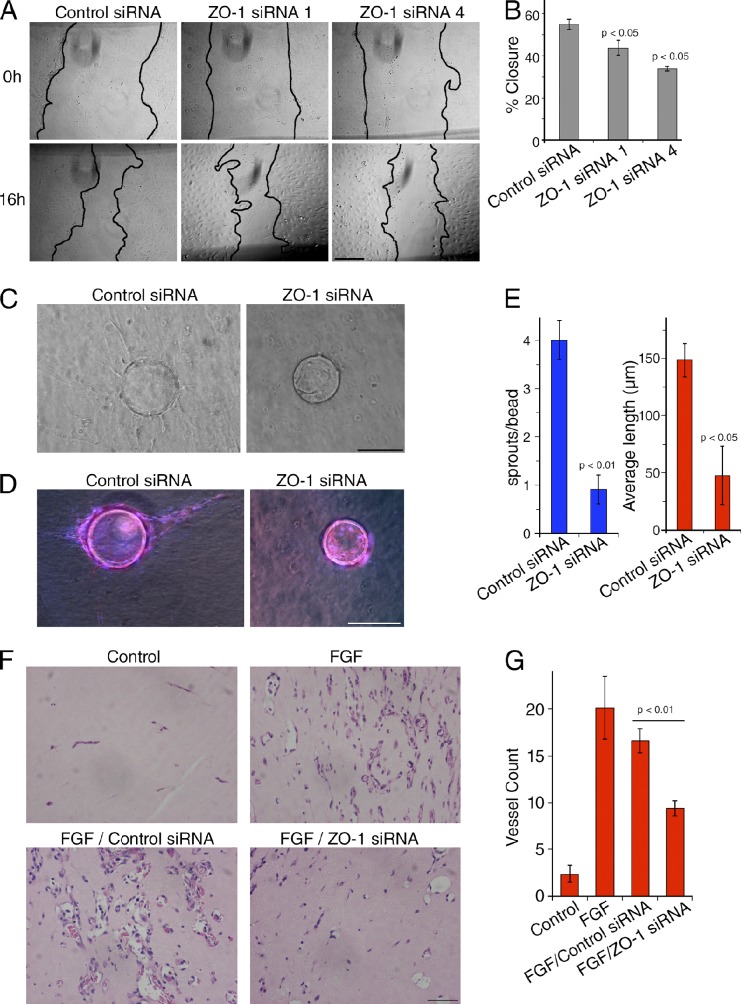
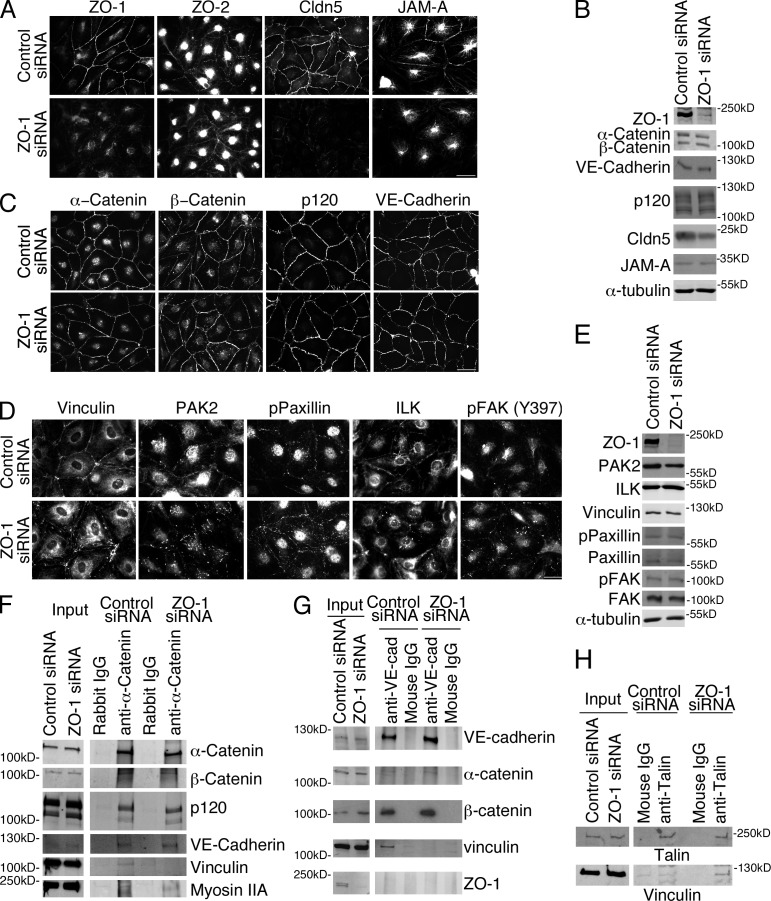
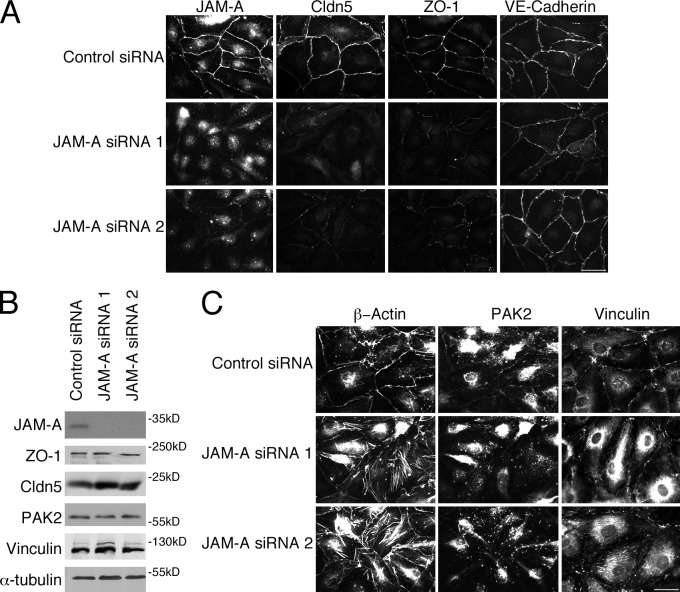
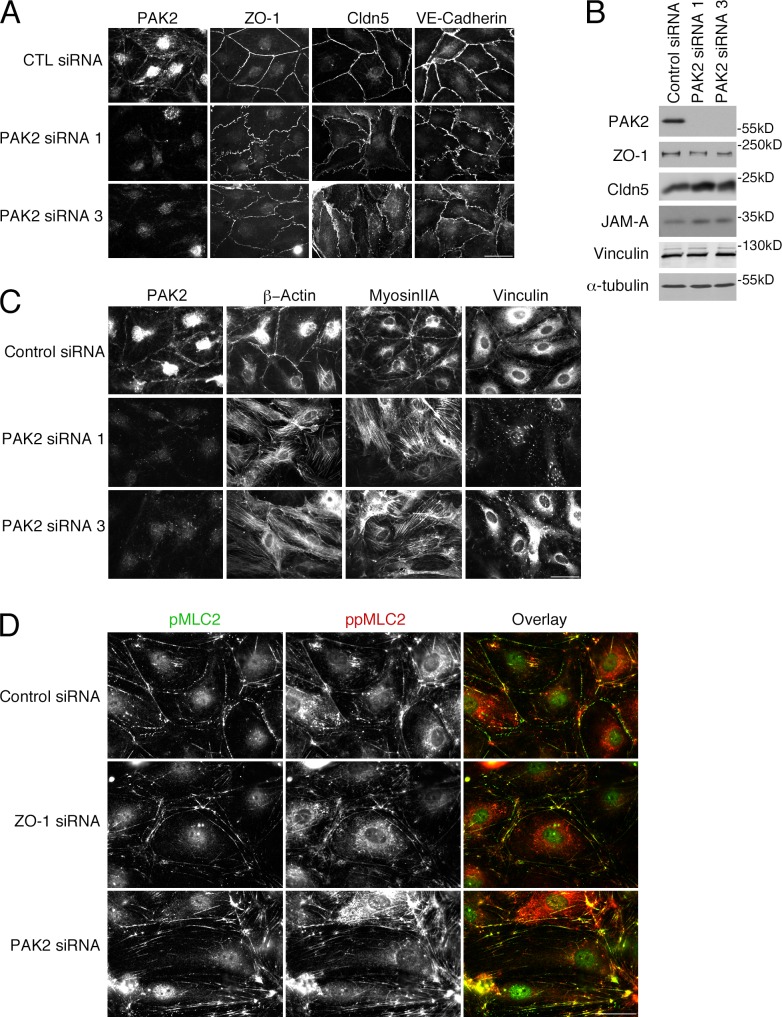
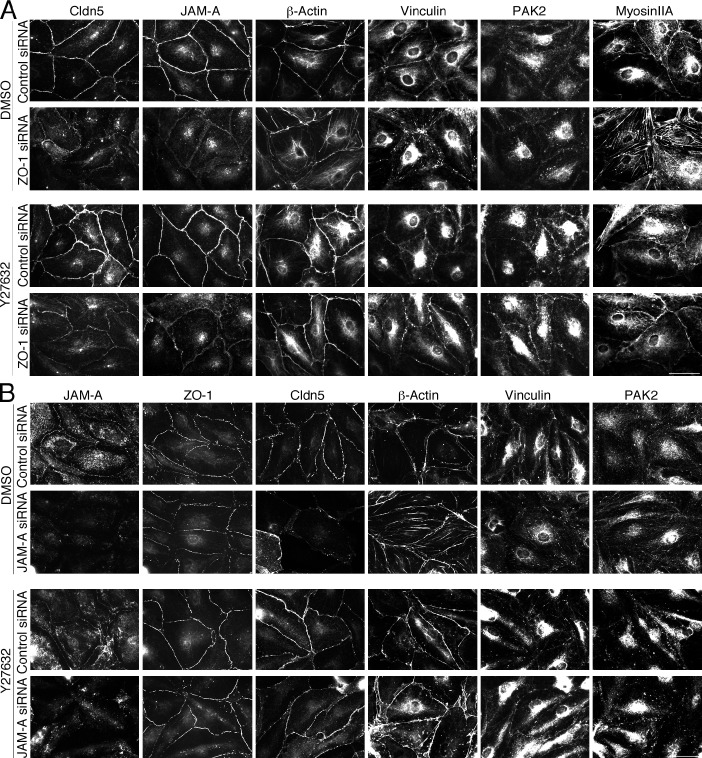
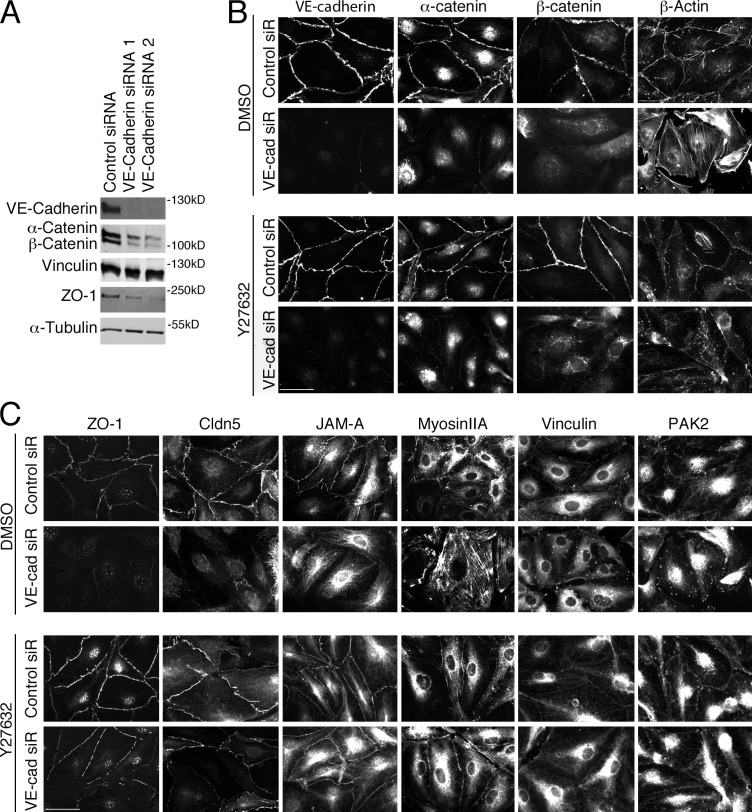
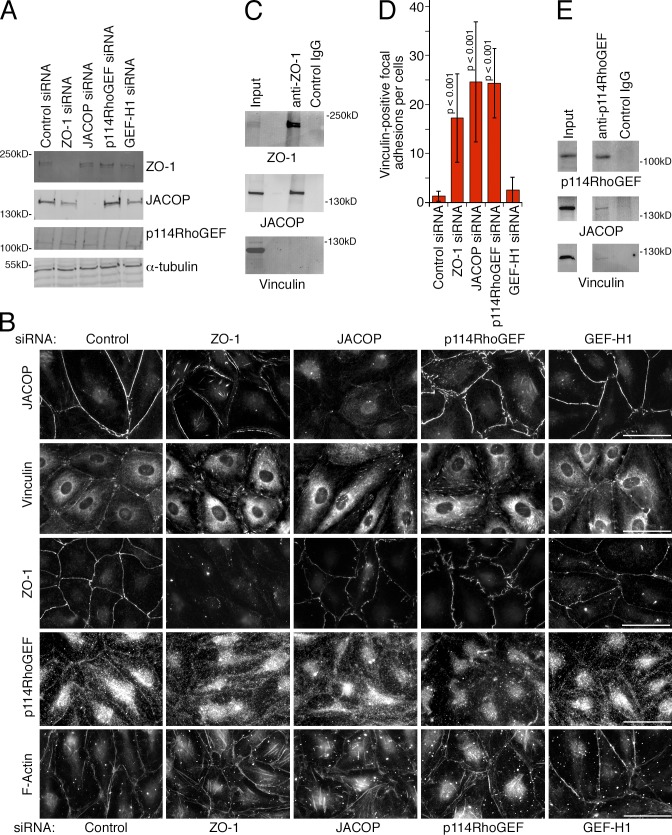
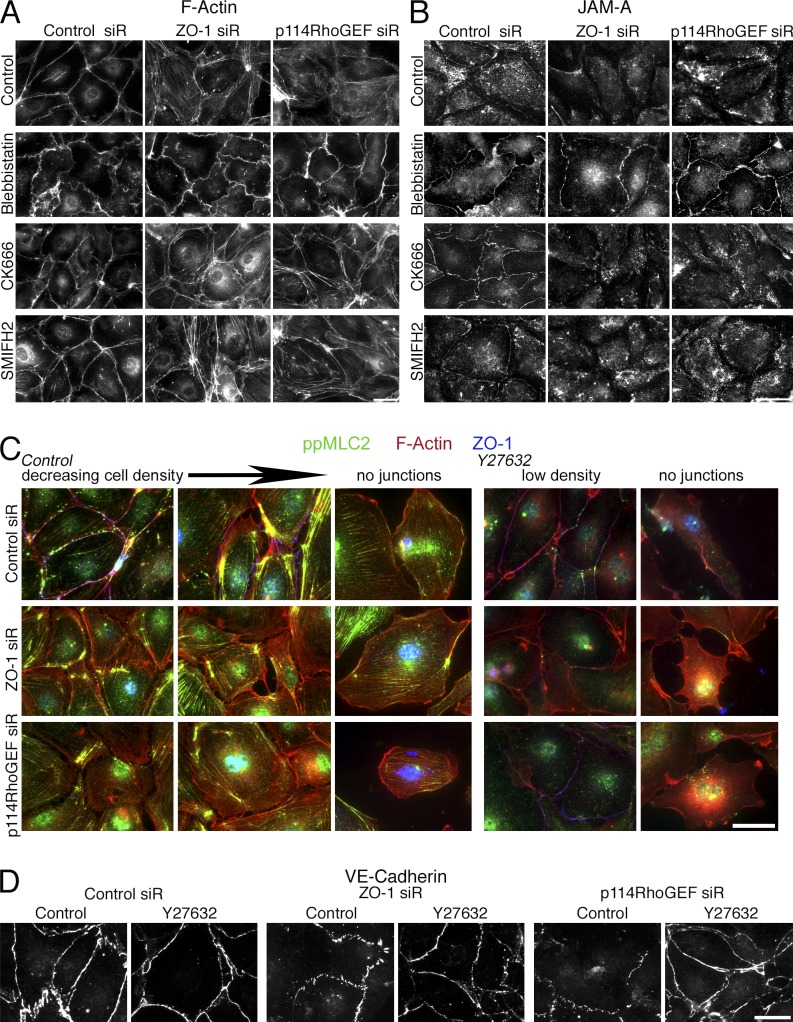
Intercellular junctions are crucial for mechanotransduction, but whether tight junctions contribute to the regulation of cell-cell tension and adherens junctions is unknown. Here, we demonstrate that the tight junction protein ZO-1 regulates tension acting on VE-cadherin-based adherens junctions, cell migration, and barrier formation of primary endothelial cells, as well as angiogenesis in vitro and in vivo. ZO-1 depletion led to tight junction disruption, redistribution of active myosin II from junctions to stress fibers, reduced tension on VE-cadherin and loss of junctional mechanotransducers such as vinculin and PAK2, and induced vinculin dissociation from the α-catenin-VE-cadherin complex. Claudin-5 depletion only mimicked ZO-1 effects on barrier formation, whereas the effects on mechanotransducers were rescued by inhibition of ROCK and phenocopied by JAM-A, JACOP, or p114RhoGEF down-regulation. ZO-1 was required for junctional recruitment of JACOP, which, in turn, recruited p114RhoGEF. ZO-1 is thus a central regulator of VE-cadherin-dependent endothelial junctions that orchestrates the spatial actomyosin organization, tuning cell-cell tension, migration, angiogenesis, and barrier formation.
© 2015 Tornavaca et al.
Figures
References
-
- Benais-Pont G., Punn A., Flores-Maldonado C., Eckert J., Raposo G., Fleming T.P., Cereijido M., Balda M.S., and Matter K.. 2003. Identification of a tight junction-associated guanine nucleotide exchange factor that activates Rho and regulates paracellular permeability. J. Cell Biol. 160:729–740 10.1083/jcb.200211047 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/J015032/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- RG/11/17/29256/BHF_/British Heart Foundation/United Kingdom
- 099173/Z/12/Z/WT_/Wellcome Trust/United Kingdom
- R01 HL75092/HL/NHLBI NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- G0900098/MRC_/Medical Research Council/United Kingdom
- R01 HL075092/HL/NHLBI NIH HHS/United States
- BB/L007584/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/H002294/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0400678/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
