

Tuning IL-2 signaling by ADP-ribosylation of CD25
- PMID: 25753532
- PMCID: PMC4354014
- DOI: 10.1038/srep08959
Tuning IL-2 signaling by ADP-ribosylation of CD25
Abstract
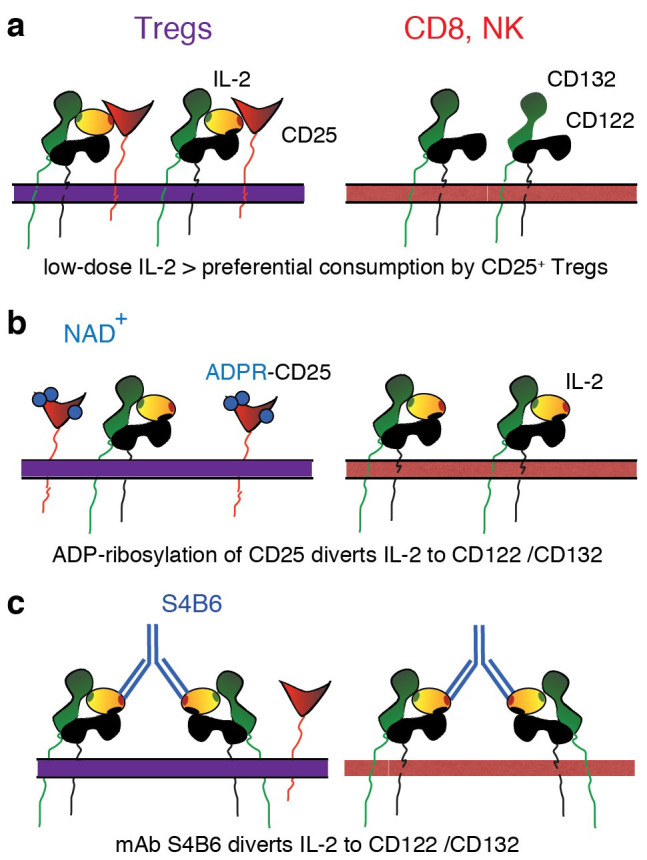
Control of immunologic tolerance and homeostasis rely on Foxp3(+)CD4(+)CD25(+) regulatory T cells (Tregs) that constitutively express the high affinity receptor for Interleukin-2, CD25. Tregs proliferate in response to injections of IL-2/anti-IL-2 antibody complexes or low doses of IL-2. However, little is known about endogenous mechanisms that regulate the sensitivity of CD25 to signaling by IL-2. Here we demonstrate that CD25 is ADP-ribosylated at Arg35 in the IL-2 binding site by ecto-ADP-ribosyltransferase ARTC2.2, a toxin-related GPI-anchored ecto-enzyme. ADP-ribosylation inhibits binding of IL-2 by CD25, IL-2- induced phosphorylation of STAT5, and IL-2-dependent cell proliferation. Our study elucidates an as-yet-unrecognized mechanism to tune IL-2 signaling. This newly found mechanism might thwart Tregs at sites of inflammation and thereby permit a more potent response of activated effector T cells.
Conflict of interest statement
F.K.-N. and F.H. receive royalties from sales of antibodies developed in the lab via MediGate GmbH, a 100% subsidiary of the University Medical Center, Hamburg.
Figures


References
-
- Sakaguchi S. Naturally arising Foxp3-expressing CD25+CD4+ regulatory T cells in immunological tolerance to self and non-self. Nat Immunol 6, 345–352 (2005). - PubMed
-
- Bluestone J. A. The yin and yang of interleukin-2-mediated immunotherapy. N Engl J Med 365, 2129–2131 (2011). - PubMed
-
- Boyman O. & Sprent J. The role of interleukin-2 during homeostasis and activation of the immune system. Nat Rev Immunol 12, 180–190 (2012). - PubMed
-
- Matko J. et al. GPI-microdomains (membrane rafts) and signaling of the multi-chain interleukin-2 receptor in human lymphoma/leukemia T cell lines. Eur J Biochem 269, 1199–1208 (2002). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
