

Redundancy in ribonucleotide excision repair: Competition, compensation, and cooperation
- PMID: 25753809
- PMCID: PMC4426009
- DOI: 10.1016/j.dnarep.2015.02.008
Redundancy in ribonucleotide excision repair: Competition, compensation, and cooperation
Abstract
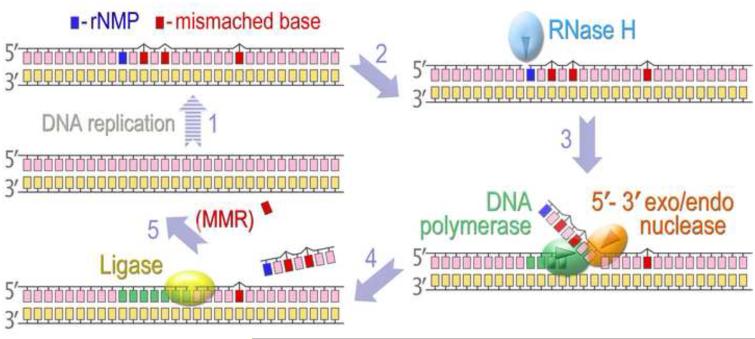
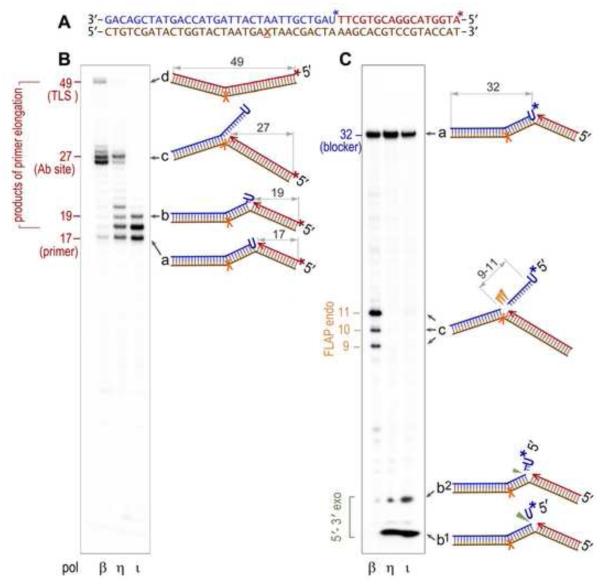
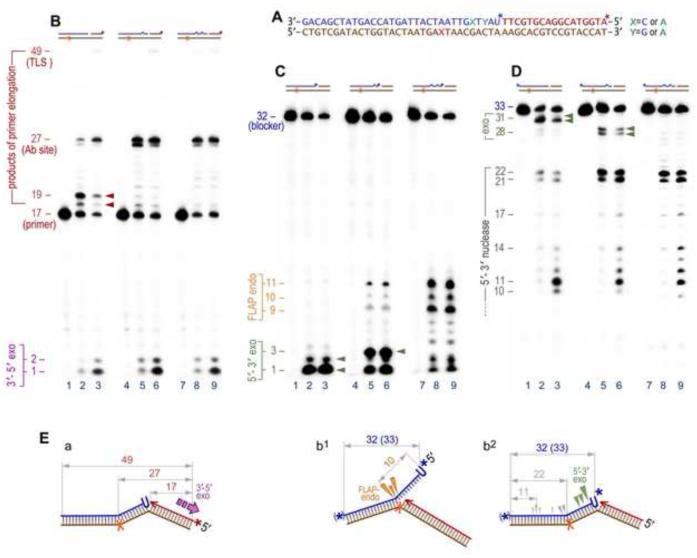
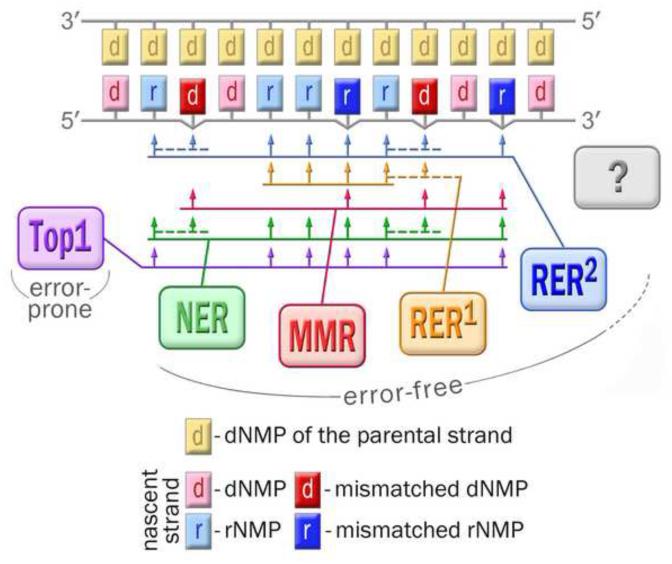
The survival of all living organisms is determined by their ability to reproduce, which in turn depends on accurate duplication of chromosomal DNA. In order to ensure the integrity of genome duplication, DNA polymerases are equipped with stringent mechanisms by which they select and insert correctly paired nucleotides with a deoxyribose sugar ring. However, this process is never 100% accurate. To fix occasional mistakes, cells have evolved highly sophisticated and often redundant mechanisms. A good example is mismatch repair (MMR), which corrects the majority of mispaired bases and which has been extensively studied for many years. On the contrary, pathways leading to the replacement of nucleotides with an incorrect sugar that is embedded in chromosomal DNA have only recently attracted significant attention. This review describes progress made during the last few years in understanding such pathways in both prokaryotes and eukaryotes. Genetic studies in Escherichia coli and Saccharomyces cerevisiae demonstrated that MMR has the capacity to replace errant ribonucleotides, but only when the base is mispaired. In contrast, the major evolutionarily conserved ribonucleotide repair pathway initiated by the ribonuclease activity of type 2 Rnase H has broad specificity. In yeast, this pathway also requires the concerted action of Fen1 and pol δ, while in bacteria it can be successfully completed by DNA polymerase I. Besides these main players, all organisms contain alternative enzymes able to accomplish the same tasks, although with differing efficiency and fidelity. Studies in bacteria have very recently demonstrated that isolated rNMPs can be removed from genomic DNA by error-free nucleotide excision repair (NER), while studies in yeast suggest the involvement of topoisomerase 1 in alternative mutagenic ribonucleotide processing. This review summarizes the most recent progress in understanding the ribonucleotide repair mechanisms in prokaryotes and eukaryotes.
Keywords: DNA polymerase I; Flap endonuclease; Mismatch repair; Nucleotide excision repair; Ribonuclease H; Ribonucleotide excision repair.
Published by Elsevier B.V.
Figures
References
-
- Vaisman A, Woodgate R. Ribonucleotide selectivity of translesion DNA synthesis polymerases. Research Signpost. 2014
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
