

FSH regulates fat accumulation and redistribution in aging through the Gαi/Ca(2+)/CREB pathway
- PMID: 25754247
- PMCID: PMC4406670
- DOI: 10.1111/acel.12331
FSH regulates fat accumulation and redistribution in aging through the Gαi/Ca(2+)/CREB pathway
Abstract
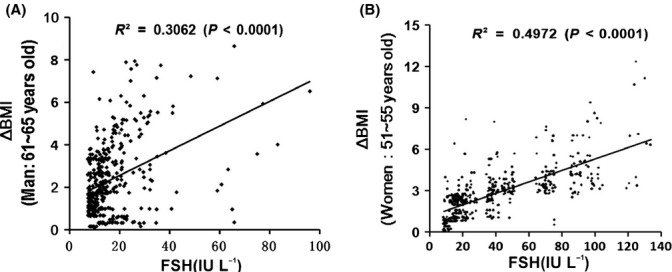
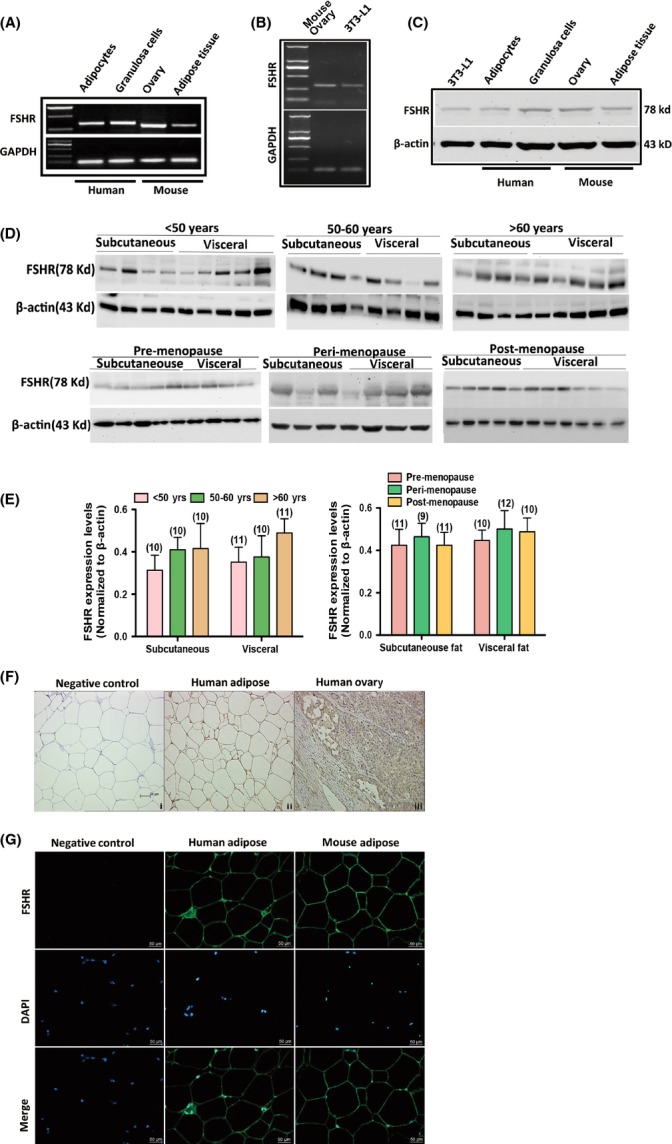
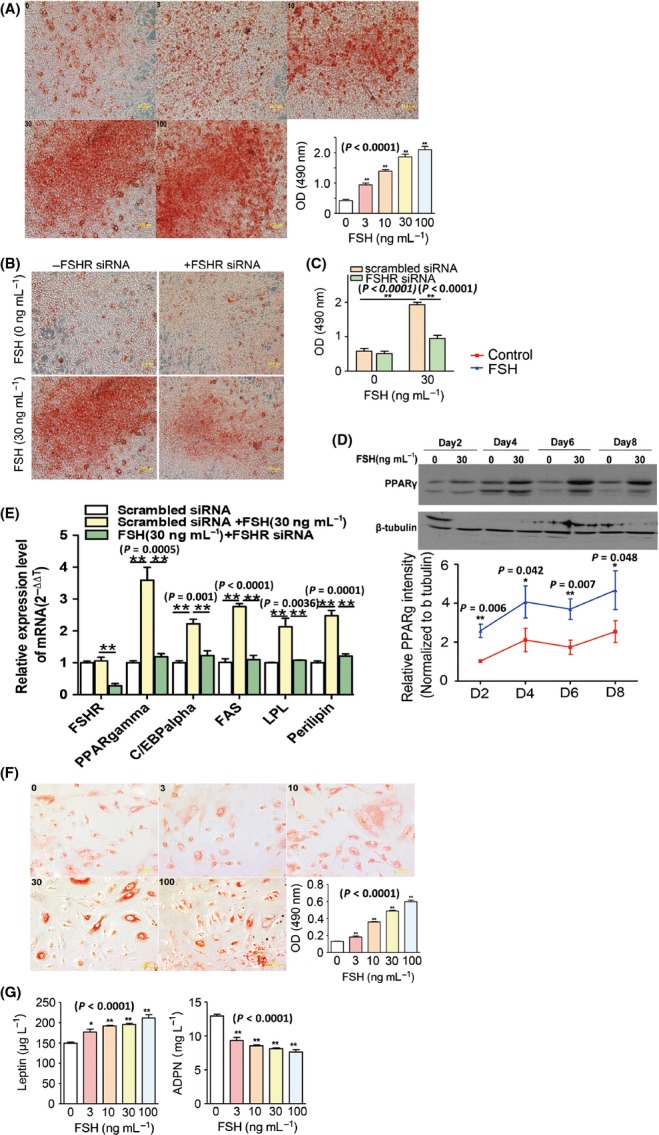
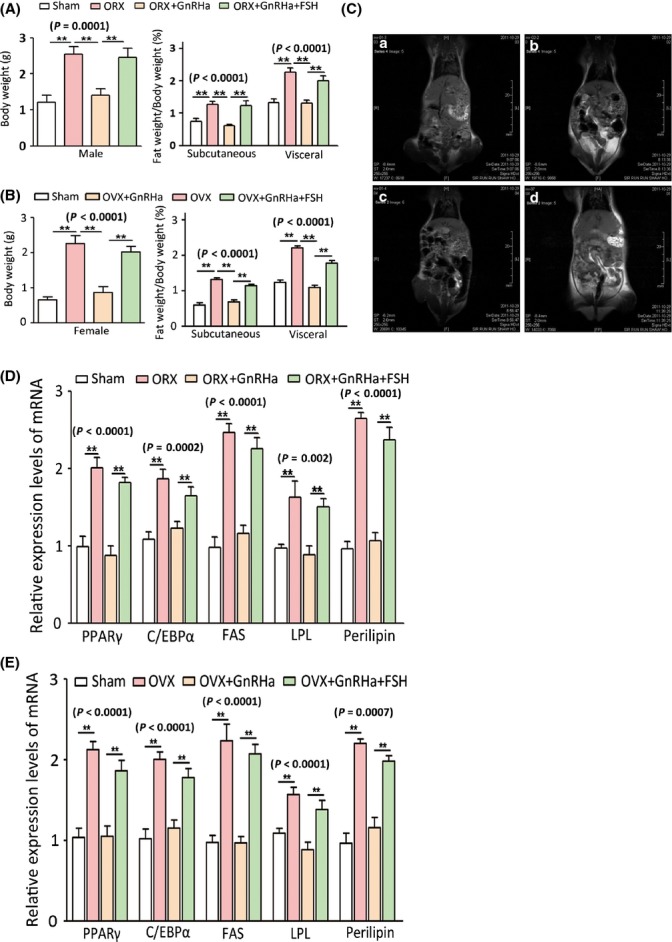
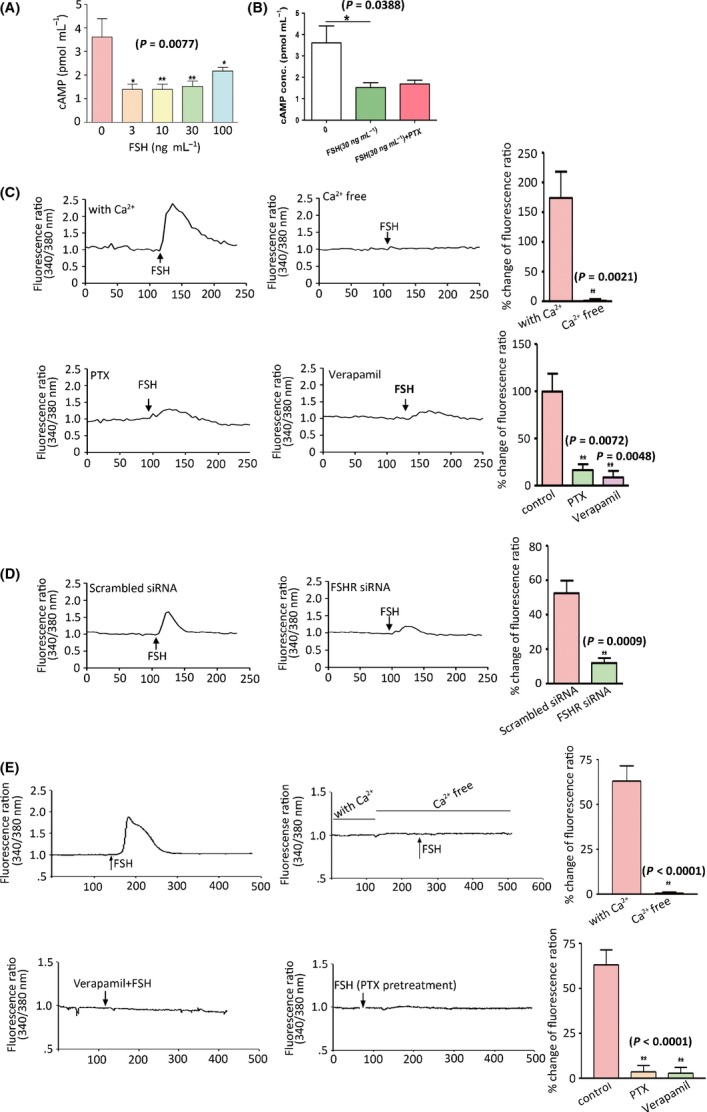
Increased fat mass and fat redistribution are commonly observed in aging populations worldwide. Although decreased circulating levels of sex hormones, androgens and oestrogens have been observed, the exact mechanism of fat accumulation and redistribution during aging remains obscure. In this study, the receptor of follicle-stimulating hormone (FSH), a gonadotropin that increases sharply and persistently with aging in both males and females, is functionally expressed in human and mouse fat tissues and adipocytes. Follicle-stimulating hormone was found to promote lipid biosynthesis and lipid droplet formation; FSH could also alter the secretion of leptin and adiponectin, but not hyperplasia, in vitro and in vivo. The effects of FSH are mediated by FSH receptors coupled to the Gαi protein; as a result, Ca(2+) influx is stimulated, cAMP-response-element-binding protein is phosphorylated, and an array of genes involved in lipid biosynthesis is activated. The present findings depict the potential of FSH receptor-mediated lipodystrophy of adipose tissues in aging. Our results also reveal the mechanism of fat accumulation and redistribution during aging of males and females.
Keywords: ageing; ca2+; endocrinology; mouse models; signal transduction; signalling.
© 2015 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Figures

References
-
- Asatiani K, Gromoll J, Eckardstein SV, Zitzmann M, Nieschlag E, Simoni M. Distribution and function of FSH receptor genetic variants in normal men. Andrologia. 2002;34:172–176. - PubMed
-
- Asayama K, Hayashibe H, Dobashi K, Uchida N, Nakane T, Kodera K, Shirahata A, Taniyama M. Decrease in serum adiponectin level due to obesity and visceral fat accumulation in children. Obes. Res. 2003;11:1072–1079. - PubMed
-
- Ausmanas MK, Tan DA, Jaisamrarn U, Tian XW, Holinka CF. Estradiol, FSH and LH profiles in nine ethnic groups of postmenopausal Asian females: the Pan-Asia Menopause (PAM) study. Climacteric. 2007;10:427–437. - PubMed
-
- Brasaemle DL. Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis. J. Lipid Res. 2007;48:2547–2559. - PubMed
-
- Cargill M, Altshuler D, Ireland J, Sklar P, Ardlie K, Patil N, Shaw N, Lane CR, Lim EP, Kalyanaraman N, Nemesh J, Ziaugra L, Friedland L, Rolfe A, Warrington J, Lipshutz R, Daley GQ, Lander ES. Characterization of single-nucleotide polymorphisms in coding regions of human genes. Nat. Genet. 1999;22:231–238. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
