

Activation of muscular TrkB by its small molecular agonist 7,8-dihydroxyflavone sex-dependently regulates energy metabolism in diet-induced obese mice
- PMID: 25754472
- PMCID: PMC4369457
- DOI: 10.1016/j.chembiol.2015.02.003
Activation of muscular TrkB by its small molecular agonist 7,8-dihydroxyflavone sex-dependently regulates energy metabolism in diet-induced obese mice
Abstract
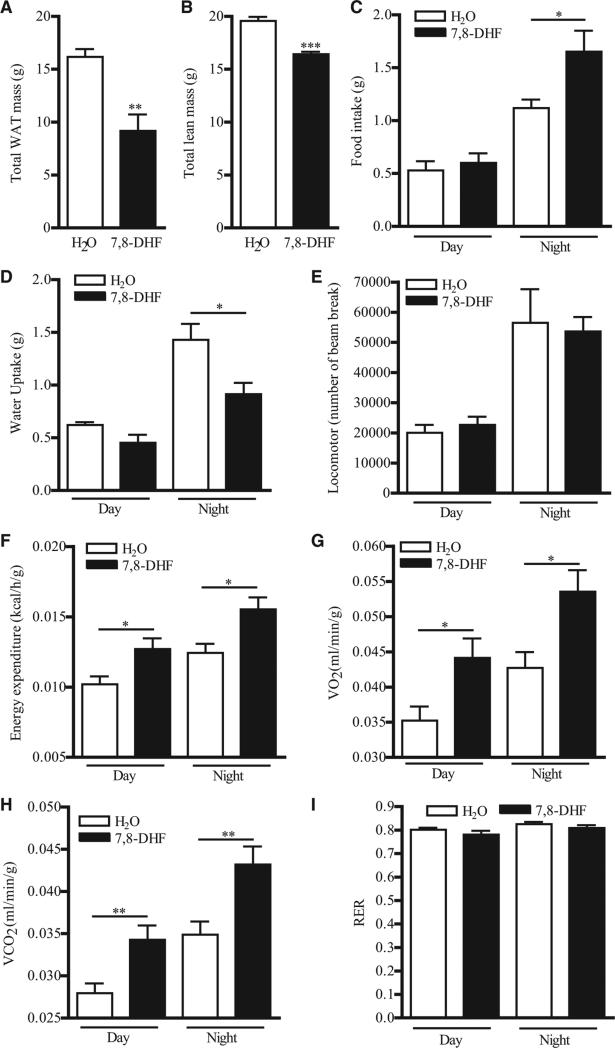
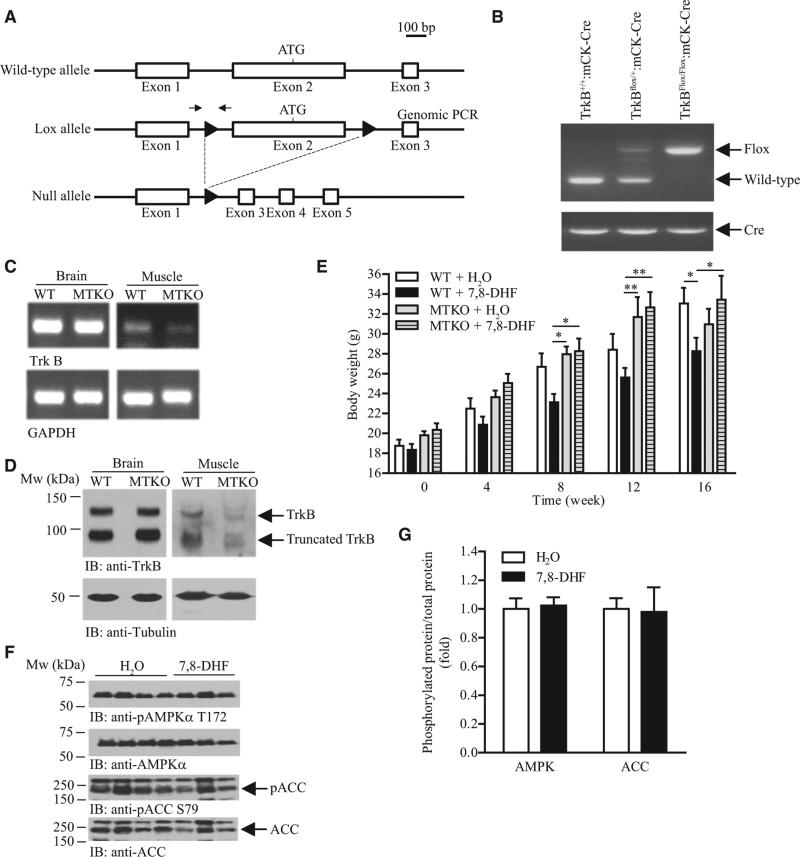
Chronic activation of brain-derived neurotrophic factor (BDNF) receptor TrkB is a potential method to prevent development of obesity, but the short half-life and nonbioavailable nature of BDNF hampers validation of the hypothesis. We report here that activation of muscular TrkB by the BDNF mimetic, 7,8-dihydroxyflavone (7,8-DHF), is sufficient to protect the development of diet-induced obesity in female mice. Using in vitro and in vivo models, we found that 7,8-DHF treatment enhanced the expression of uncoupling protein 1 (UCP1) and AMP-activated protein kinase (AMPK) activity in skeletal muscle, which resulted in increased systemic energy expenditure, reduced adiposity, and improved insulin sensitivity in female mice fed a high-fat diet. This antiobesity activity of 7,8-DHF is muscular TrkB-dependent as 7,8-DHF cannot mitigate diet-induced obesity in female muscle-specific TrkB knockout mice. Hence, our data reveal that chronic activation of muscular TrkB is useful in alleviating obesity and its complications.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures





Comment in
-
Fueling up skeletal muscle to reduce obesity: A TrkB story.Chem Biol. 2015 Mar 19;22(3):311-2. doi: 10.1016/j.chembiol.2015.03.004. Chem Biol. 2015. PMID: 25794433 Free PMC article.
References
-
- Andero R, Daviu N, Escorihuela RM, Nadal R, Armario A. 7,8-Dihydroxyflavone, a TrkB receptor agonist, blocks long-term spatial memory impairment caused by immobilization stress in rats. Hippocampus. 2012;22:399–408. - PubMed
-
- Ayala JE, Bracy DP, McGuinness OP, Wasserman DH. Considerations in the design of hyperinsulinemic-euglycemic clamps in the conscious mouse. Diabetes. 2006;55:390–397. - PubMed
-
- Baker-Andresen D, Flavell CR, Li X, Bredy TW. Activation of BDNF signaling prevents the return of fear in female mice. Learn. Mem. 2013;20:237–240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
