

Pentraxins coordinate excitatory synapse maturation and circuit integration of parvalbumin interneurons
- PMID: 25754824
- PMCID: PMC4368480
- DOI: 10.1016/j.neuron.2015.02.020
Pentraxins coordinate excitatory synapse maturation and circuit integration of parvalbumin interneurons
Erratum in
-
Pentraxins Coordinate Excitatory Synapse Maturation and Circuit Integration of Parvalbumin Interneurons.Neuron. 2016 May 4;90(3):661. doi: 10.1016/j.neuron.2016.04.024. Neuron. 2016. PMID: 27151642 Free PMC article. No abstract available.
Abstract
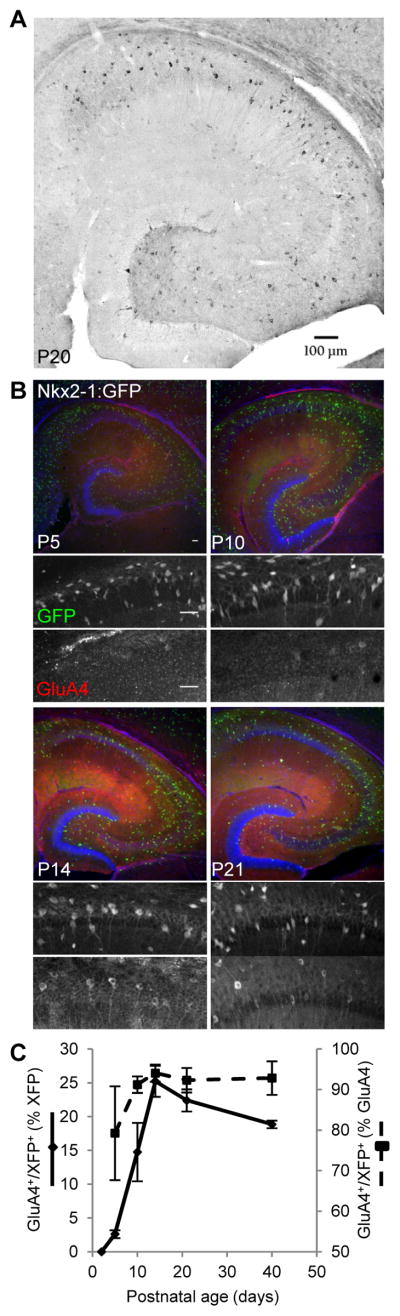
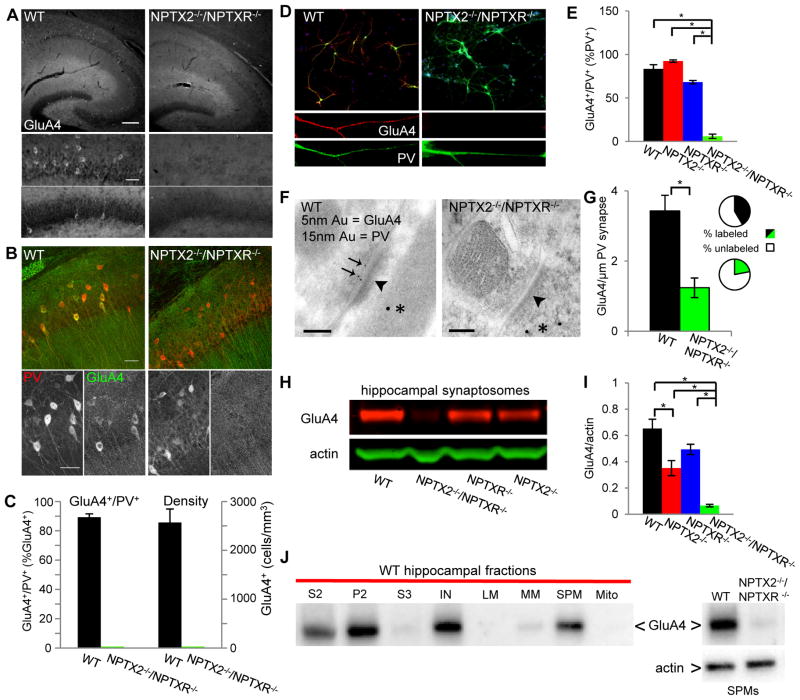
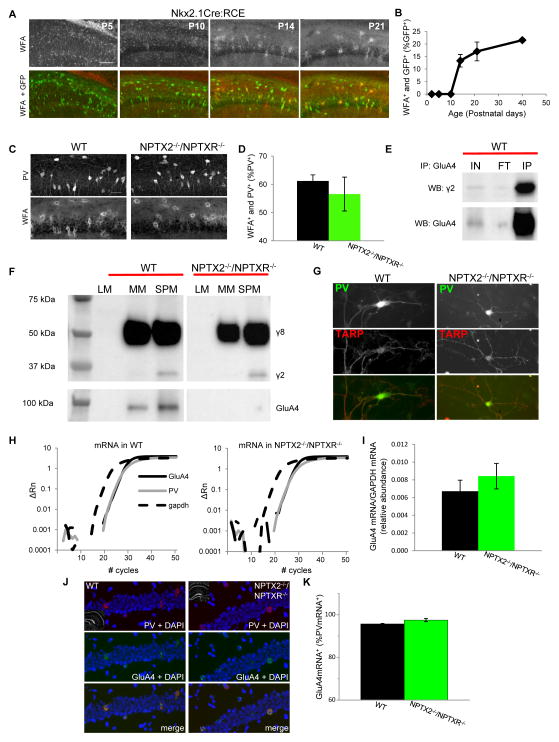
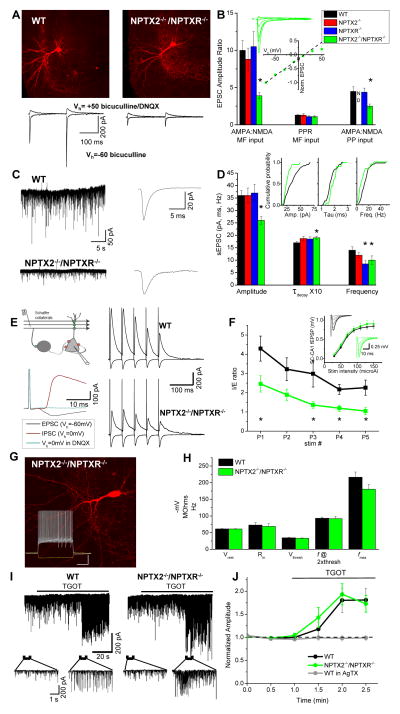
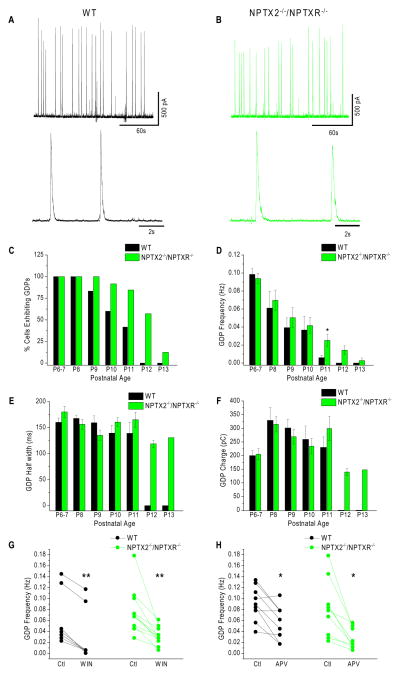
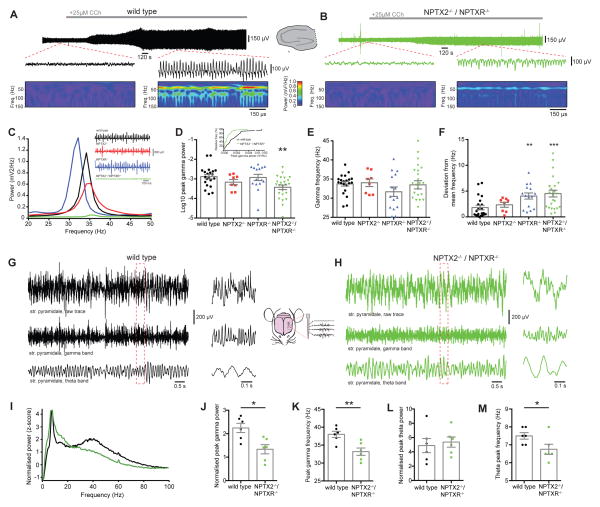
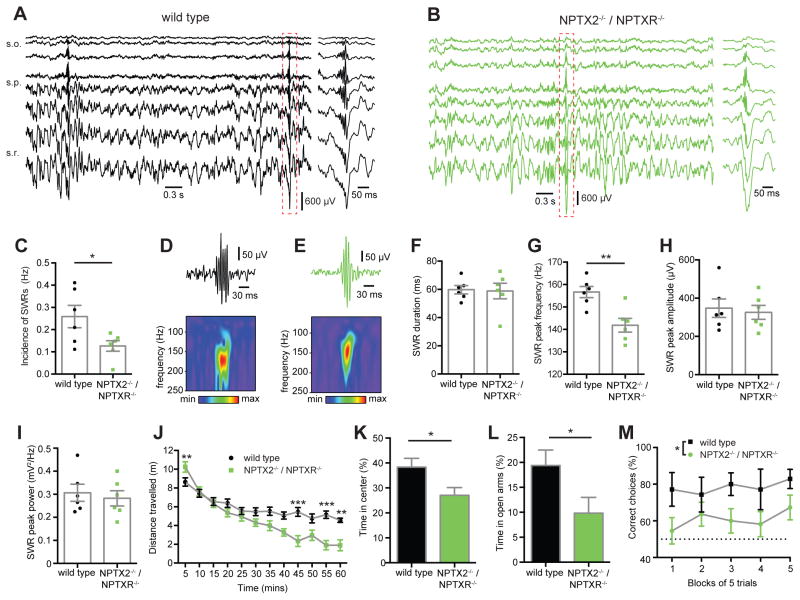
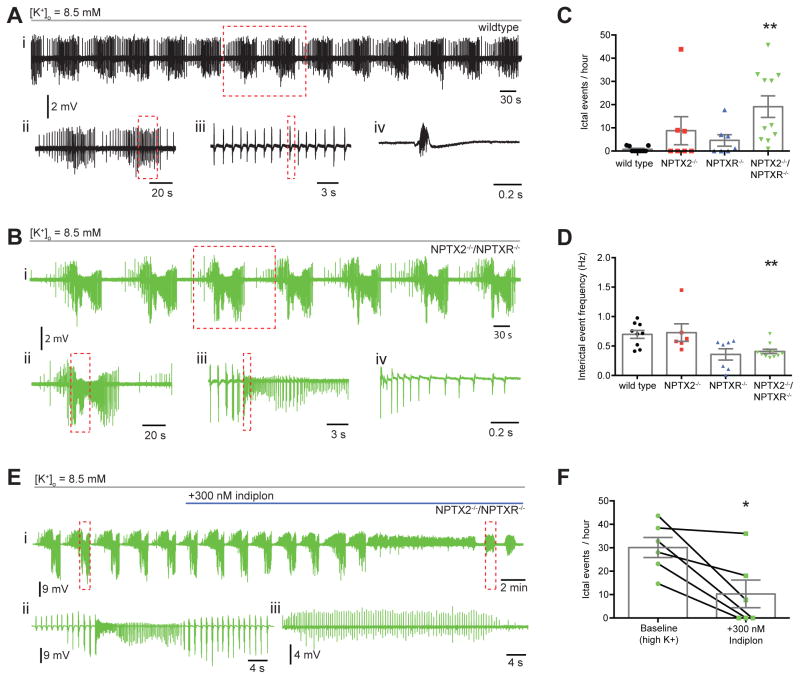
Circuit computation requires precision in the timing, extent, and synchrony of principal cell (PC) firing that is largely enforced by parvalbumin-expressing, fast-spiking interneurons (PVFSIs). To reliably coordinate network activity, PVFSIs exhibit specialized synaptic and membrane properties that promote efficient afferent recruitment such as expression of high-conductance, rapidly gating, GluA4-containing AMPA receptors (AMPARs). We found that PVFSIs upregulate GluA4 during the second postnatal week coincident with increases in the AMPAR clustering proteins NPTX2 and NPTXR. Moreover, GluA4 is dramatically reduced in NPTX2(-/-)/NPTXR(-/-) mice with consequent reductions in PVFSI AMPAR function. Early postnatal NPTX2(-/-)/NPTXR(-/-) mice exhibit delayed circuit maturation with a prolonged critical period permissive for giant depolarizing potentials. Juvenile NPTX2(-/-)/NPTXR(-/-) mice display reduced feedforward inhibition yielding a circuit deficient in rhythmogenesis and prone to epileptiform discharges. Our findings demonstrate an essential role for NPTXs in controlling network dynamics highlighting potential therapeutic targets for disorders with inhibition/excitation imbalances such as schizophrenia.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Barad Z, Shevtsova O, Arbuthnott GW, Leitch B. Selective loss of AMPA receptors at corticothalamic synapses in the epileptic stargazer mouse. Neuroscience. 2012;217:19–31. - PubMed
-
- Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat Rev Neurosci. 2007;8:45–56. - PubMed
-
- Ben-Ari Y, Gaiarsa JL, Tyzio R, Khazipov R. GABA: a pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol Rev. 2007;87:1215–1284. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
