

Mechanosensitive control of plant growth: bearing the load, sensing, transducing, and responding
- PMID: 25755656
- PMCID: PMC4337334
- DOI: 10.3389/fpls.2015.00052
Mechanosensitive control of plant growth: bearing the load, sensing, transducing, and responding
Abstract
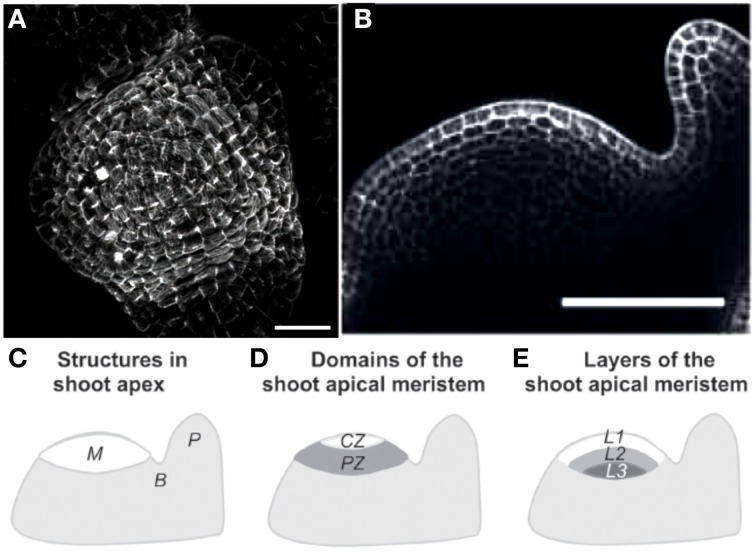
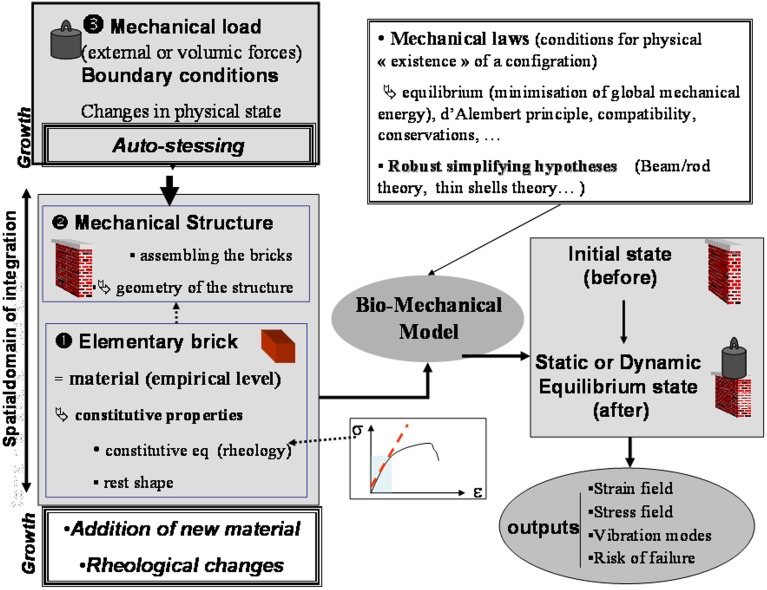
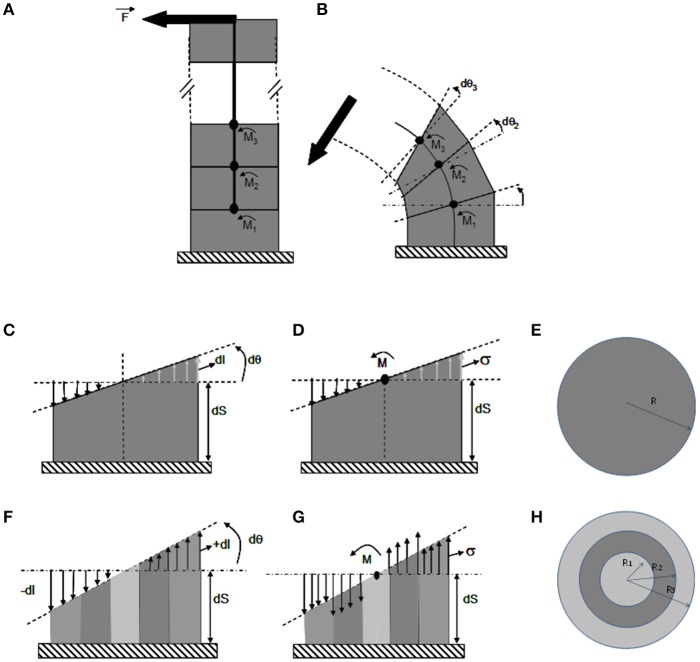
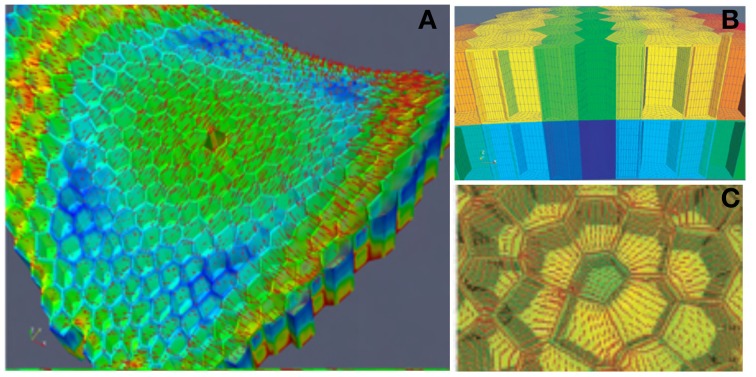
As land plants grow and develop, they encounter complex mechanical challenges, especially from winds and turgor pressure. Mechanosensitive control over growth and morphogenesis is an adaptive trait, reducing the risks of breakage or explosion. This control has been mostly studied through experiments with artificial mechanical loads, often focusing on cellular or molecular mechanotransduction pathway. However, some important aspects of mechanosensing are often neglected. (i) What are the mechanical characteristics of different loads and how are loads distributed within different organs? (ii) What is the relevant mechanical stimulus in the cell? Is it stress, strain, or energy? (iii) How do mechanosensing cells signal to meristematic cells? Without answers to these questions we cannot make progress analyzing the mechanobiological effects of plant size, plant shape, tissue distribution and stiffness, or the magnitude of stimuli. This situation is rapidly changing however, as systems mechanobiology is being developed, using specific biomechanical and/or mechanobiological models. These models are instrumental in comparing loads and responses between experiments and make it possible to quantitatively test biological hypotheses describing the mechanotransduction networks. This review is designed for a general plant science audience and aims to help biologists master the models they need for mechanobiological studies. Analysis and modeling is broken down into four steps looking at how the structure bears the load, how the distributed load is sensed, how the mechanical signal is transduced, and then how the plant responds through growth. Throughout, two examples of adaptive responses are used to illustrate this approach: the thigmorphogenetic syndrome of plant shoots bending and the mechanosensitive control of shoot apical meristem (SAM) morphogenesis. Overall this should provide a generic understanding of systems mechanobiology at work.
Keywords: biomechanics; curvature; mechanobiology; mechanotransduction; stress; thigmomorphogenesis; turgor pressure; wind.
Figures



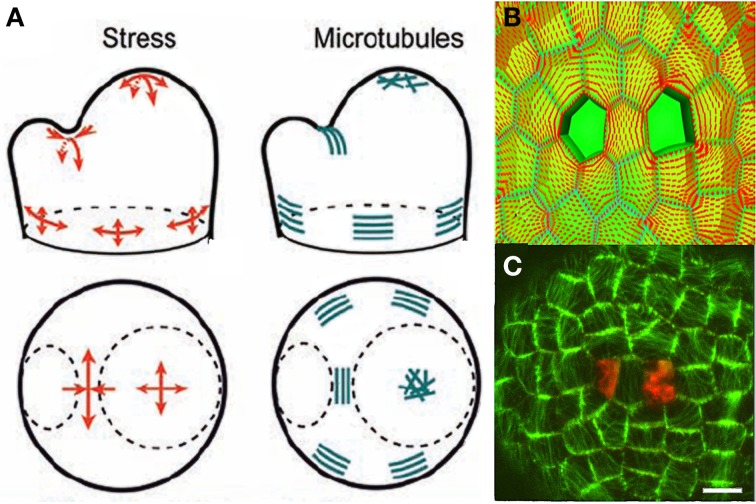
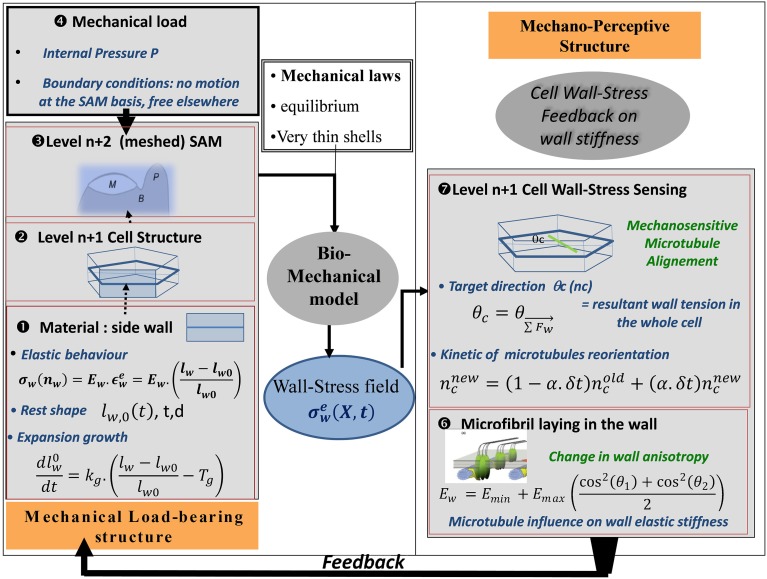

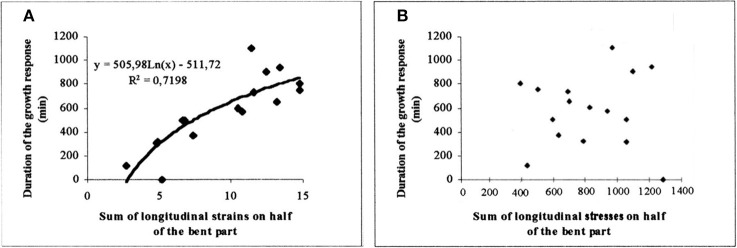

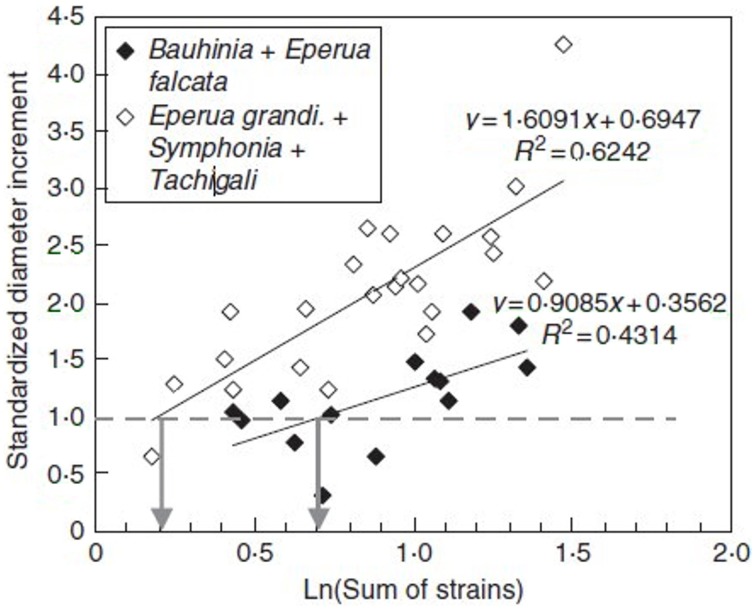
References
-
- Barbacci A., Diener J., Hemon P., Adam B., Dones N., Reveret L., et al. (2014). A robust videogrametric method for the velocimetry of wind-induced motion in trees. Agri. Forest Meteorol. 184, 220–229 10.1016/j.agrformet.2013.10.003 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials