

Polyol specificity of recombinant Arabidopsis thaliana sorbitol dehydrogenase studied by enzyme kinetics and in silico modeling
- PMID: 25755662
- PMCID: PMC4337239
- DOI: 10.3389/fpls.2015.00091
Polyol specificity of recombinant Arabidopsis thaliana sorbitol dehydrogenase studied by enzyme kinetics and in silico modeling
Abstract
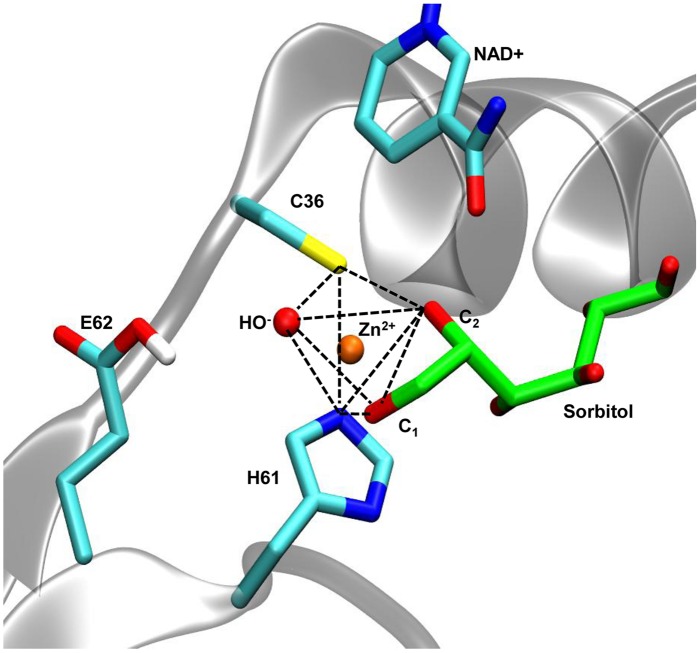
Polyols are enzymatically-produced plant compounds which can act as compatible solutes during periods of abiotic stress. Nicotinamide adenine dinucleotide(+)-dependent SORBITOL DEHYDROGENASE (SDH, E. C. 1.1.1.14) from Arabidopsis thaliana L. sorbitol dehydrogenase (AtSDH) is capable of oxidizing several polyols including sorbitol, ribitol, and xylitol. In the present study, enzymatic assays using recombinant AtSDH demonstrated a higher specificity constant for xylitol compared to sorbitol and ribitol, all of which are C2 (S) and C4 (R) polyols. Enzyme activity was reduced by preincubation with ethylenediaminetetraacetic acid, indicating a requirement for zinc ions. In humans, it has been proposed that sorbitol becomes part of a pentahedric coordination sphere of the catalytic zinc during the reaction mechanism. In order to determine the validity of this pentahedric coordination model in a plant SDH, homology modeling, and Molecular Dynamics simulations of AtSDH ternary complexes with the three polyols were performed using crystal structures of human and Bemisia argentifolii (Genn.) (Hemiptera: Aleyrodidae) SDHs as scaffolds. The results indicate that the differences in interaction with structural water molecules correlate very well with the observed enzymatic parameters, validate the proposed pentahedric coordination of the catalytic zinc ion in a plant SDH, and provide an explanation for why AtSDH shows a preference for polyols with a chirality of C2 (S) and C4 (R).
Keywords: Arabidopsis thaliana; homology modeling; molecular dynamics simulation; pentavalent zinc; polyol.
Figures





References
-
- Bais P., Moon S. M., He K., Leitao R., Dreher K., Walk T., et al. (2010). PlantMetabolomics.org: a web portal for plant metabolomics experiments. Plant Physiol. 152 1807–1816 10.1104/pp.109.151027 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous