

Evaluation of the synuclein-γ (SNCG) gene as a PPARγ target in murine adipocytes, dorsal root ganglia somatosensory neurons, and human adipose tissue
- PMID: 25756178
- PMCID: PMC4355072
- DOI: 10.1371/journal.pone.0115830
Evaluation of the synuclein-γ (SNCG) gene as a PPARγ target in murine adipocytes, dorsal root ganglia somatosensory neurons, and human adipose tissue
Abstract
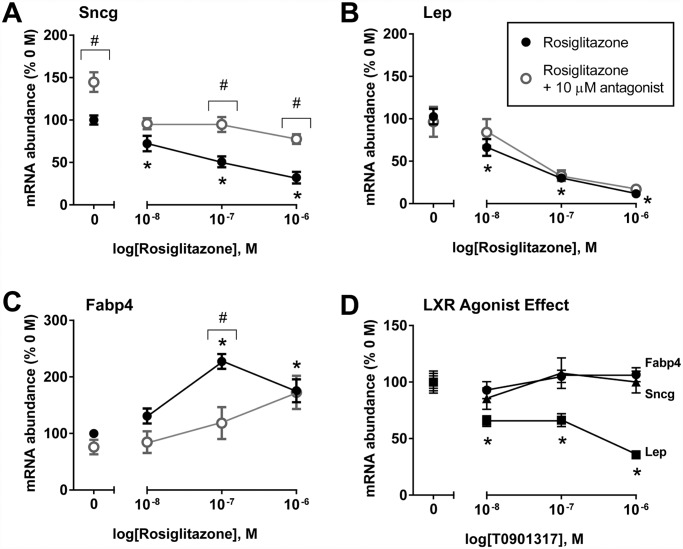
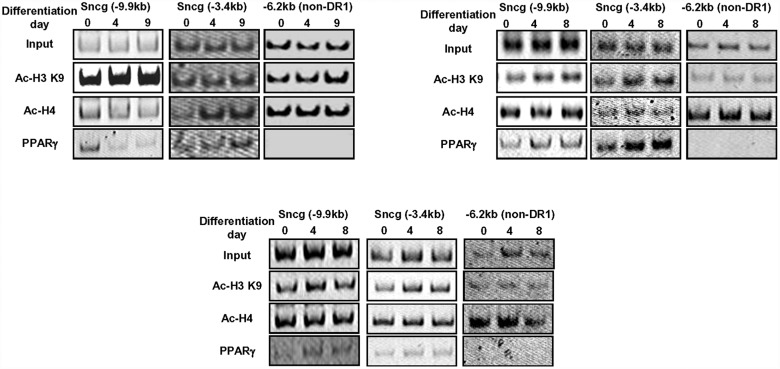
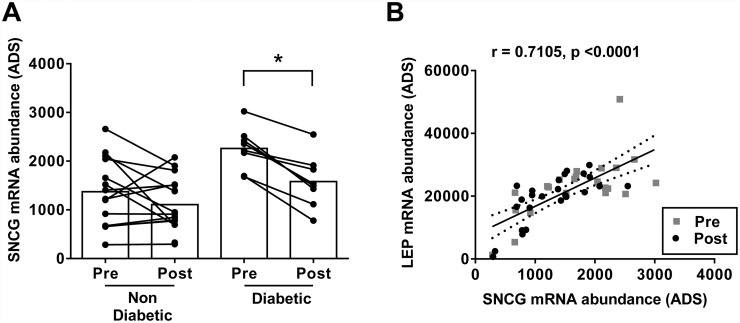
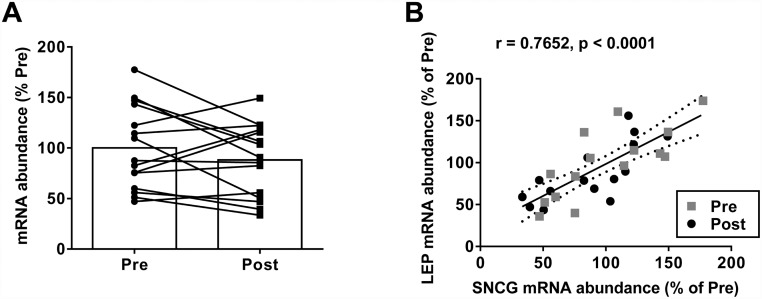
Recent evidence in adipocytes points to a role for synuclein-γ in metabolism and lipid droplet dynamics, but interestingly this factor is also robustly expressed in peripheral neurons. Specific regulation of the synuclein-γ gene (Sncg) by PPARγ requires further evaluation, especially in peripheral neurons, prompting us to test if Sncg is a bona fide PPARγ target in murine adipocytes and peripheral somatosensory neurons derived from the dorsal root ganglia (DRG). Sncg mRNA was decreased in 3T3-L1 adipocytes (~68%) by rosiglitazone, and this effect was diminished by the PPARγ antagonist T0070907. Chromatin immunoprecipitation experiments confirmed PPARγ protein binding at two promoter sequences of Sncg during 3T3-L1 adipogenesis. Rosiglitazone did not affect Sncg mRNA expression in murine cultured DRG neurons. In subcutaneous human WAT samples from two cohorts treated with pioglitazone (>11 wks), SNCG mRNA expression was reduced, albeit highly variable and most evident in type 2 diabetes. Leptin (Lep) expression, thought to be coordinately-regulated with Sncg based on correlations in human adipose tissue, was also reduced in 3T3-L1 adipocytes by rosiglitazone. However, Lep was unaffected by PPARγ antagonist, and the LXR agonist T0901317 significantly reduced Lep expression (~64%) while not impacting Sncg. The results support the concept that synuclein-γ shares some, but not all, gene regulators with leptin and is a PPARγ target in adipocytes but not DRG neurons. Regulation of synuclein-γ by cues such as PPARγ agonism in adipocytes is logical based on recent evidence for an important role for synuclein-γ in the maintenance and dynamics of adipocyte lipid droplets.
Conflict of interest statement
Figures
References
-
- van Baak MA (2001) The peripheral sympathetic nervous system in human obesity. Obes Rev 2: 3–14. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
