

Innate immune response to Streptococcus pyogenes depends on the combined activation of TLR13 and TLR2
- PMID: 25756897
- PMCID: PMC4355416
- DOI: 10.1371/journal.pone.0119727
Innate immune response to Streptococcus pyogenes depends on the combined activation of TLR13 and TLR2
Abstract
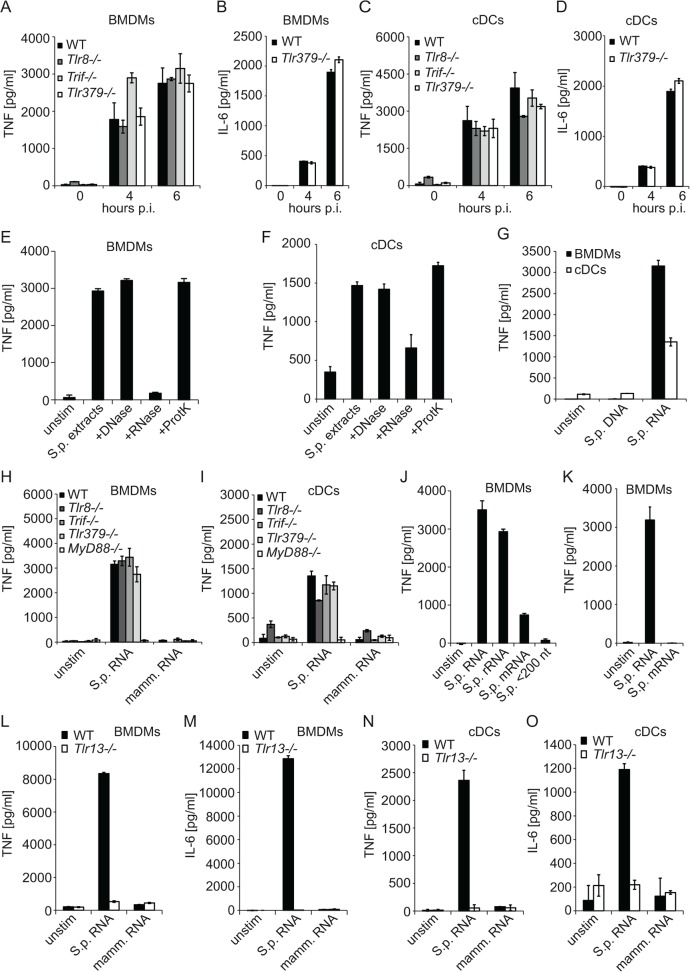
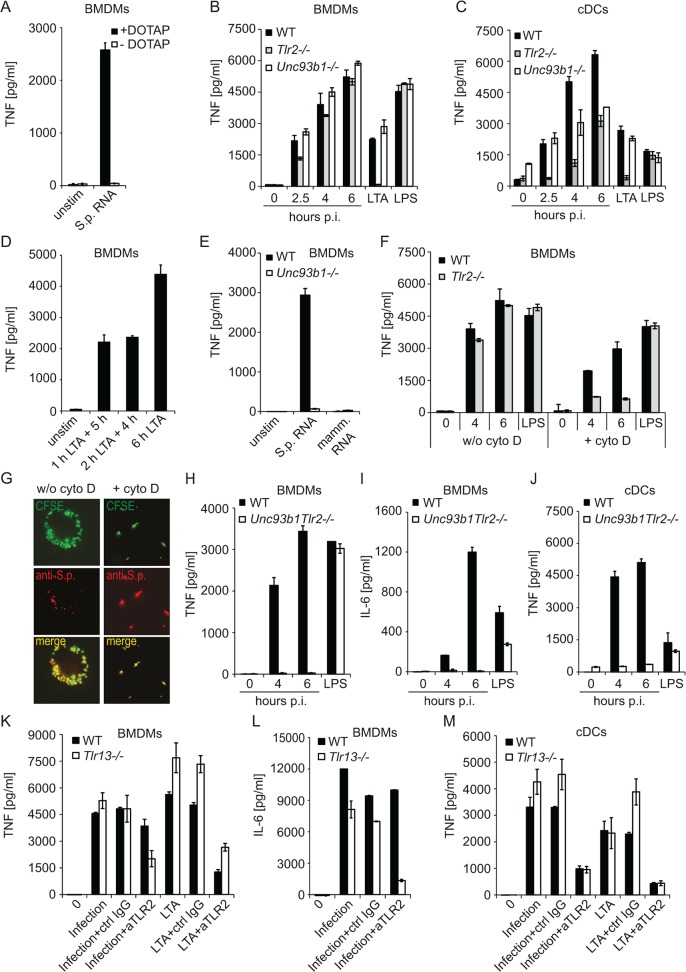
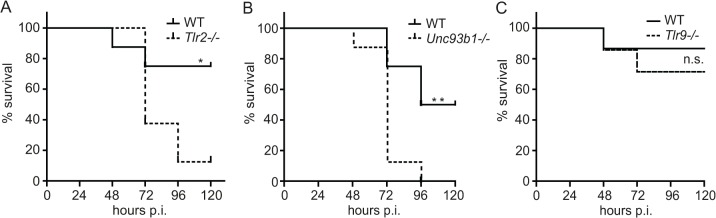
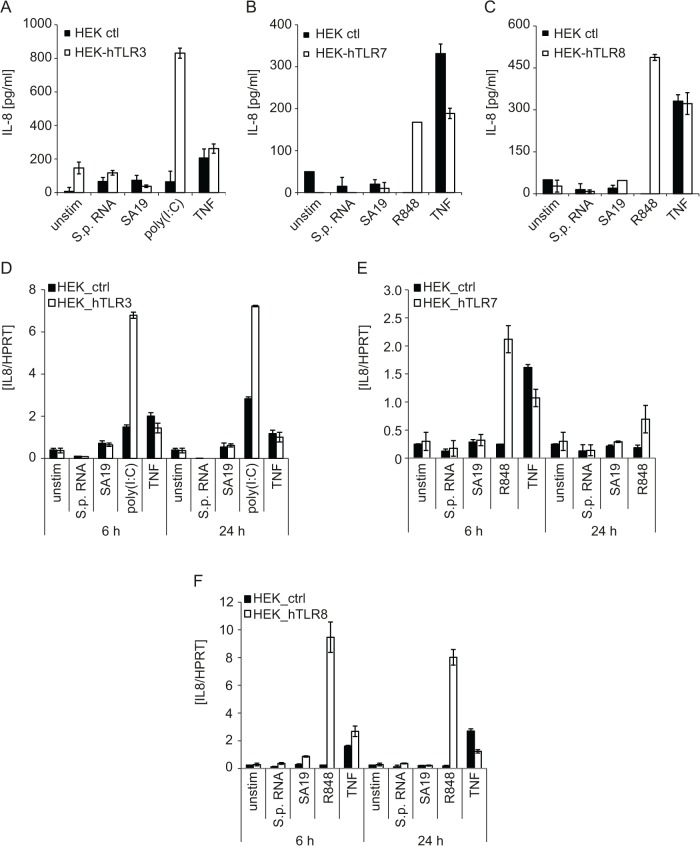
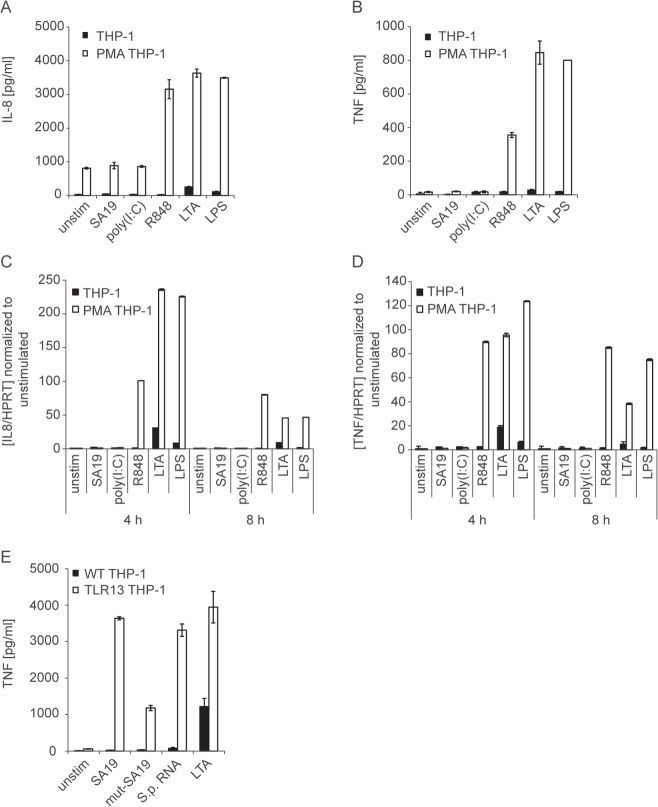
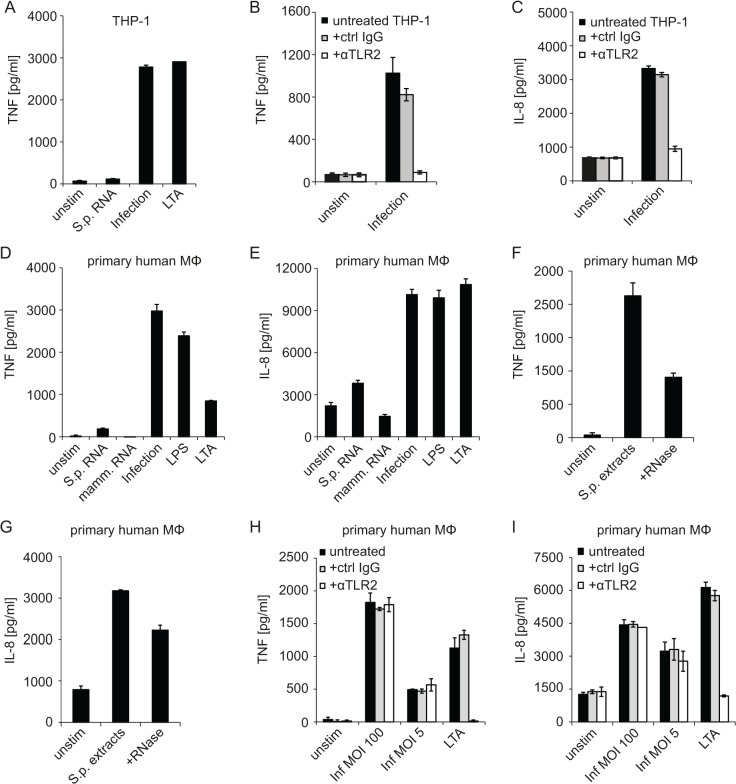
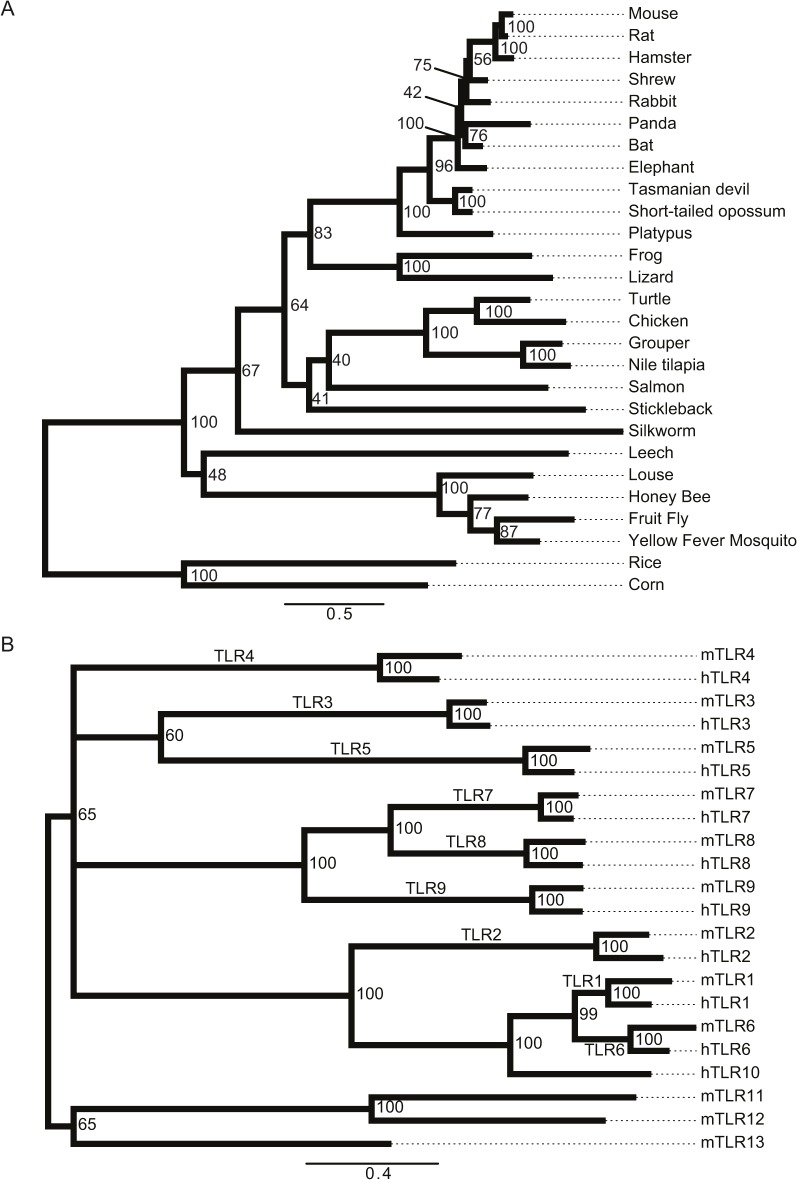
Innate immune recognition of the major human-specific Gram-positive pathogen Streptococcus pyogenes is not understood. Here we show that mice employ Toll-like receptor (TLR) 2- and TLR13-mediated recognition of S. pyogenes. These TLR pathways are non-redundant in the in vivo context of animal infection, but are largely redundant in vitro, as only inactivation of both of them abolishes inflammatory cytokine production by macrophages and dendritic cells infected with S. pyogenes. Mechanistically, S. pyogenes is initially recognized in a phagocytosis-independent manner by TLR2 and subsequently by TLR13 upon internalization. We show that the TLR13 response is specifically triggered by S. pyogenes rRNA and that Tlr13-/- cells respond to S. pyogenes infection solely by engagement of TLR2. TLR13 is absent from humans and, remarkably, we find no equivalent route for S. pyogenes RNA recognition in human macrophages. Phylogenetic analysis reveals that TLR13 occurs in all kingdoms but only in few mammals, including mice and rats, which are naturally resistant against S. pyogenes. Our study establishes that the dissimilar expression of TLR13 in mice and humans has functional consequences for recognition of S. pyogenes in these organisms.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
