

CXCR2 modulates bone marrow vascular repair and haematopoietic recovery post-transplant
- PMID: 25757087
- PMCID: PMC4654909
- DOI: 10.1111/bjh.13335
CXCR2 modulates bone marrow vascular repair and haematopoietic recovery post-transplant
Abstract
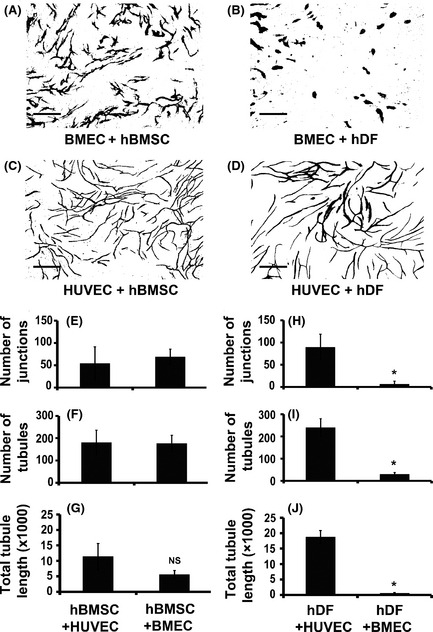
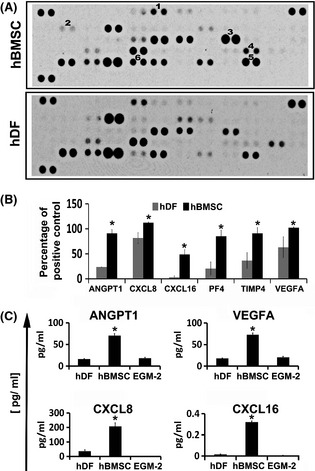
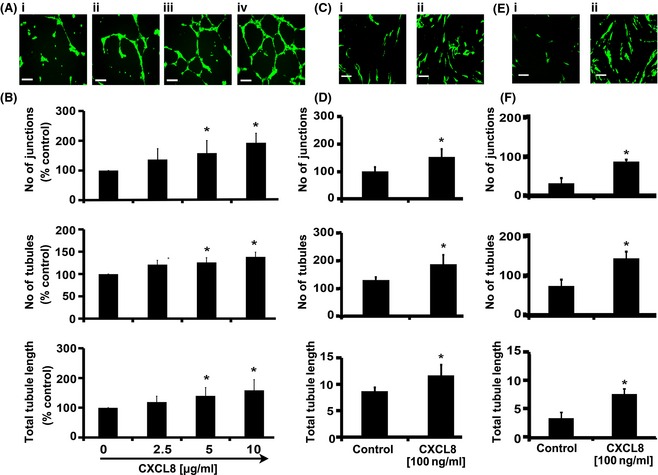
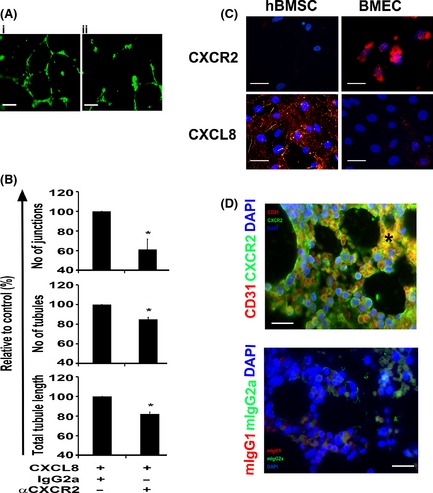
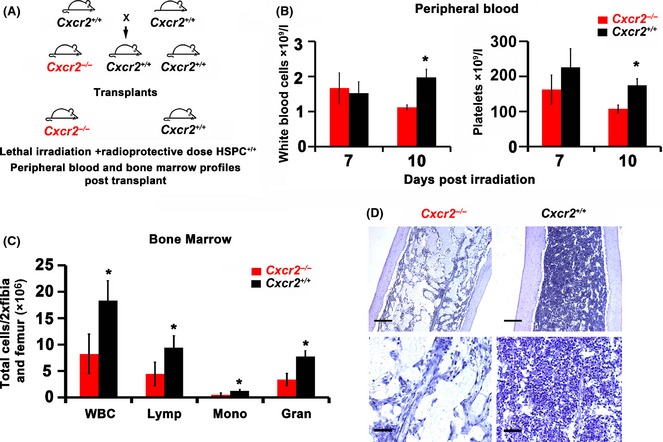
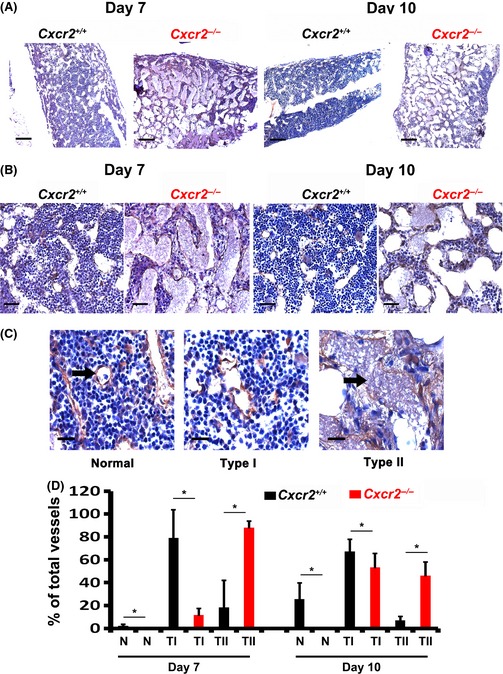
Murine models of bone marrow transplantation show that pre-conditioning regimens affect the integrity of the bone marrow endothelium and that the repair of this vascular niche is an essential pre-requisite for successful haematopoietic stem and progenitor cell engraftment. Little is known about the angiogenic pathways that play a role in the repair of the human bone marrow vascular niche. We therefore established an in vitro humanized model, composed of bone marrow stromal and endothelial cells and have identified several pro-angiogenic factors, VEGFA, ANGPT1, CXCL8 and CXCL16, produced by the stromal component of this niche. We demonstrate for the first time that addition of CXCL8 or inhibition of its receptor, CXCR2, modulates blood vessel formation in our bone marrow endothelial niche model. Compared to wild type, Cxcr2(-/-) mice displayed a reduction in bone marrow cellularity and delayed platelet and leucocyte recovery following myeloablation and bone marrow transplantation. The delay in bone marrow recovery correlated with impaired bone marrow vascular repair. Taken together, our data demonstrate that CXCR2 regulates bone marrow blood vessel repair/regeneration and haematopoietic recovery, and clinically may be a therapeutic target for improving bone marrow transplantation.
Keywords: CXCL8; CXCR2; bone marrow transplantation; bone marrow vascular niche; stem cell niche.
© 2015 The Authors. British Journal of Haematology published by John Wiley & Sons Ltd.
Figures
References
-
- Avecilla, S.T. , Hattori, K. , Heissig, B. , Tejada, R. , Liao, F. , Shido, K. , Jin, D.K. , Dias, S. , Zhang, F. , Hartman, T.E. , Hackett, N.R. , Crystal, R.G. , Witte, L. , Hicklin, D.J. , Bohlen, P. , Eaton, D. , Lyden, D. , de Sauvage, F. & Rafii, S. (2004) Chemokine‐mediated interaction of hematopoietic progenitors with the bone marrow vascular niche is required for thrombopoiesis. Nature Medicine, 10, 64–71. - PubMed
-
- Barker, J.E. (1994) Sl/Sld hematopoietic progenitors are deficient in situ. Experimental Hematology, 22, 174–177. - PubMed
-
- Barker, J.E. (1997) Early transplantation to a normal microenvironment prevents the development of Steel hematopoietic stem cell defects. Experimental Hematology, 25, 542–547. - PubMed
-
- de Barros, A.P. , Takiya, C.M. , Garzoni, L.R. , Leal‐Ferreira, M.L. , Dutra, H.S. , Chiarini, L.B. , Meirelles, M.N. , Borojevic, R. & Rossi, M.I. (2010) Osteoblasts and bone marrow mesenchymal stromal cells control hematopoietic stem cell migration and proliferation in 3D in vitro model. PLoS ONE, 5, e9093. - PMC - PubMed
-
- Bikfalvi, A. & Han, Z.C. (1994) Angiogenic factors are hematopoietic growth factors and vice versa. Leukemia, 8, 523–529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
