

Cell density sensing alters TGF-β signaling in a cell-type-specific manner, independent from Hippo pathway activation
- PMID: 25758862
- PMCID: PMC4437188
- DOI: 10.1016/j.devcel.2015.01.011
Cell density sensing alters TGF-β signaling in a cell-type-specific manner, independent from Hippo pathway activation
Abstract
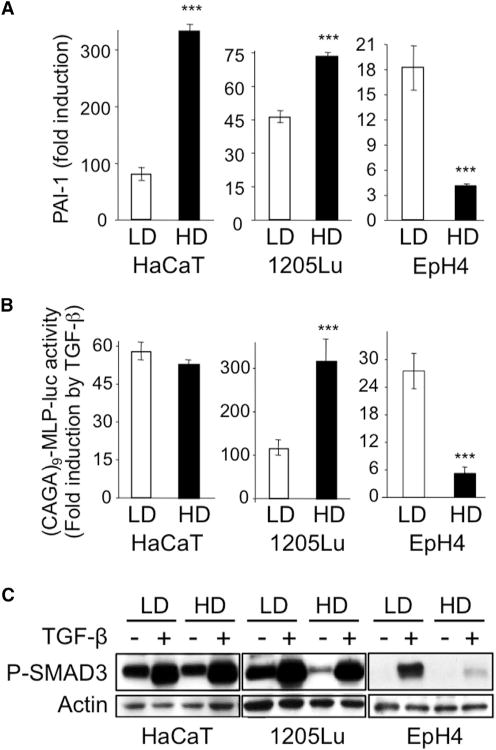
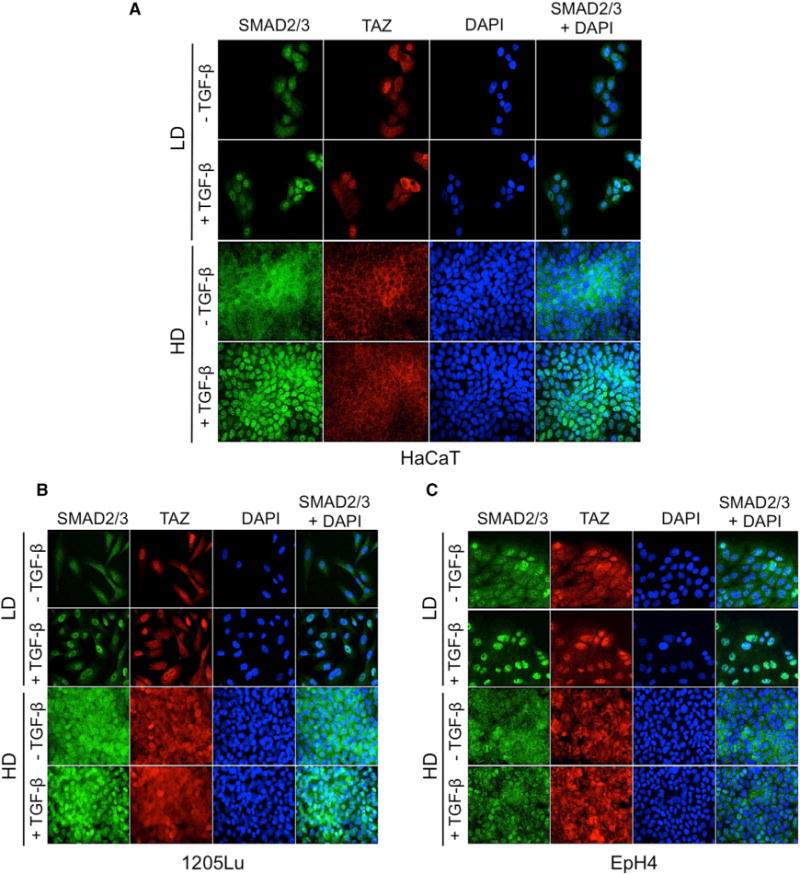
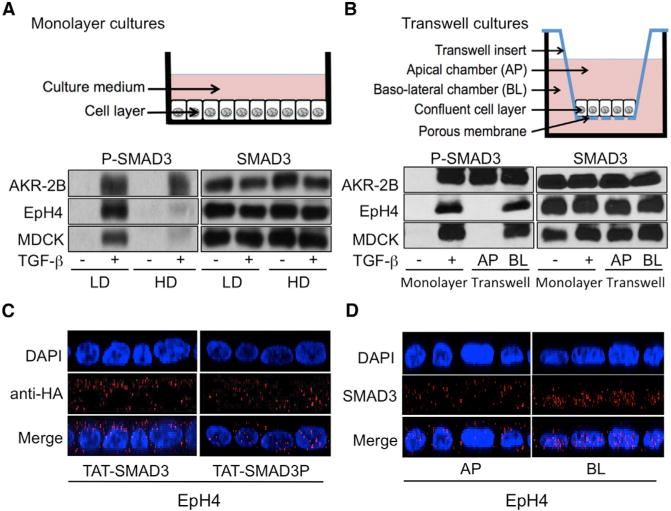
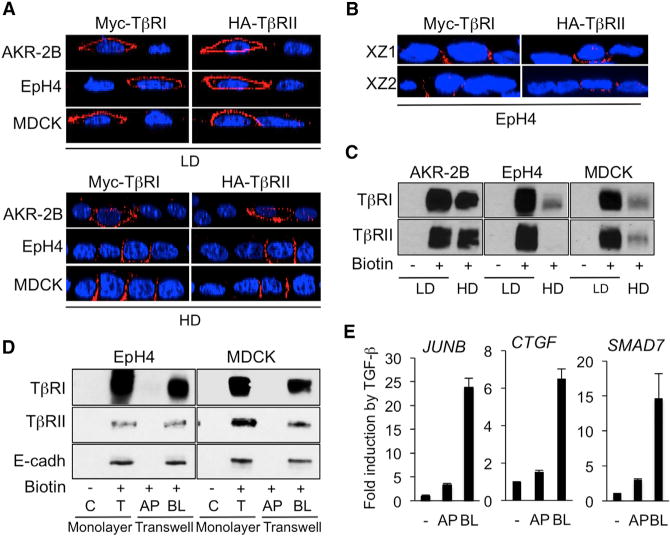
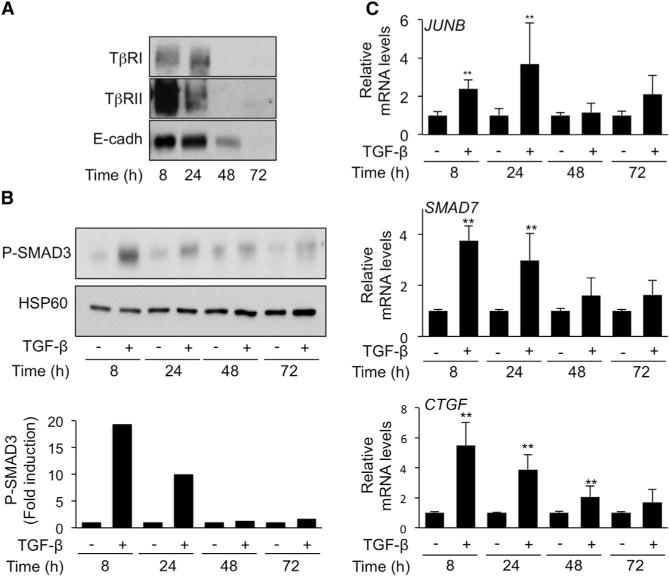
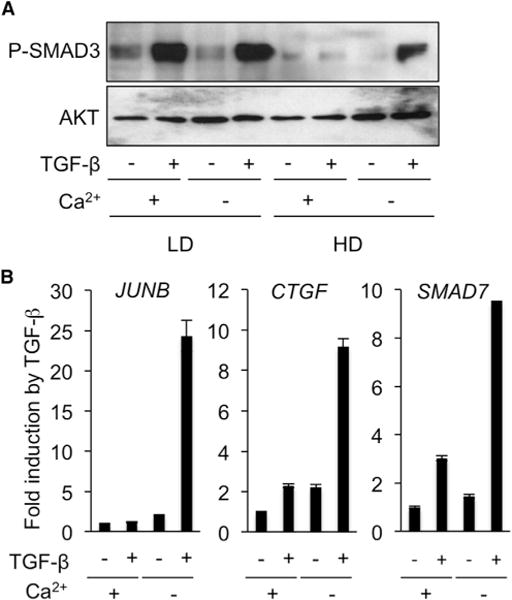
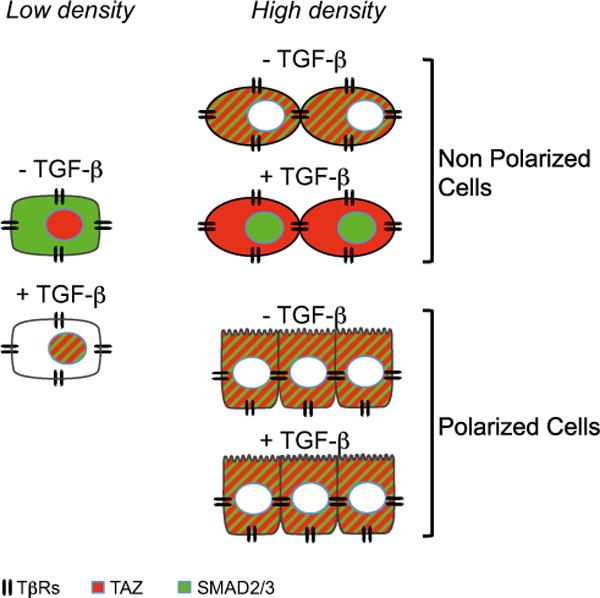
Cell-cell contacts inhibit cell growth and proliferation in part by activating the Hippo pathway that drives the phosphorylation and nuclear exclusion of the transcriptional coactivators YAP and TAZ. Cell density and Hippo signaling have also been reported to block transforming growth factor β (TGF-β) responses, based on the ability of phospho-YAP/TAZ to sequester TGF-β-activated SMAD complexes in the cytoplasm. Herein, we provide evidence that epithelial cell polarization interferes with TGF-β signaling well upstream and independent of cytoplasmic YAP/TAZ. Rather, polarized basolateral presentation of TGF-β receptors I and II deprives apically delivered TGF-β of access to its receptors. Basolateral ligand delivery nonetheless remains entirely effective to induce TGF-β responses. These data demonstrate that cell-type-specific inhibition of TGF-β signaling by cell density is restricted to polarized epithelial cells and reflects the polarized distribution of TGF-β receptors, which thus affects SMAD activation irrespective of Hippo pathway activation.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Alexaki VI, Javelaud D, Mauviel A. JNK supports survival in melanoma cells by controlling cell cycle arrest and apoptosis. Pigment Cell Melanoma Res. 2008;21:429–438. - PubMed
-
- Beyer TA, Weiss A, Khomchuk Y, Huang K, Ogunjimi AA, Varelas X, Wrana JL. Switch enhancers interpret TGF-β and Hippo signaling to control cell fate in human embryonic stem cells. Cell Rep. 2013;5:1611–1624. - PubMed
-
- Chen X, Xu H, Yuan P, Fang F, Huss M, Vega VB, Wong E, Orlov YL, Zhang W, Jiang J, et al. Integration of external signaling pathways with the core transcriptional network in embryonic stem cells. Cell. 2008;133:1106–1117. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
