

Vaccination with adjuvanted recombinant neuraminidase induces broad heterologous, but not heterosubtypic, cross-protection against influenza virus infection in mice
- PMID: 25759506
- PMCID: PMC4453582
- DOI: 10.1128/mBio.02556-14
Vaccination with adjuvanted recombinant neuraminidase induces broad heterologous, but not heterosubtypic, cross-protection against influenza virus infection in mice
Abstract
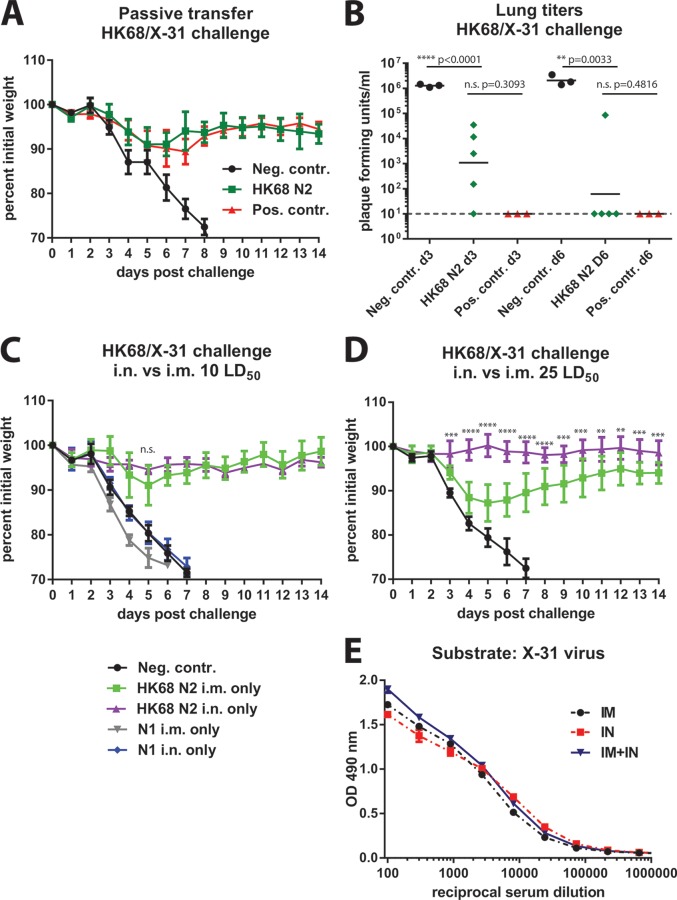
In an attempt to assess the cross-protective potential of the influenza virus neuraminidase (NA) as a vaccine antigen, different subtypes of recombinant NA were expressed in a baculovirus system and used to vaccinate mice prior to lethal challenge with homologous, heterologous, or heterosubtypic viruses. Mice immunized with NA of subtype N2 were completely protected from morbidity and mortality in a homologous challenge and displayed significantly reduced viral lung titers. Heterologous challenge with a drifted strain resulted in morbidity but no mortality. Similar results were obtained for challenge experiments with N1 NA. Mice immunized with influenza B virus NA (from B/Yamagata/16/88) displayed no morbidity when sublethally infected with the homologous strain and, importantly, were completely protected from morbidity and mortality when lethally challenged with the prototype Victoria lineage strain or a more recent Victoria lineage isolate. Upon analyzing the NA content in 4 different inactivated-virus vaccine formulations from the 2013-2014 season via Western blot assay and enzyme-linked immunosorbent assay quantification, we found that the amount of NA does indeed vary across vaccine brands. We also measured hemagglutinin (HA) and NA endpoint titers in pre- and postvaccination human serum samples from individuals who received a trivalent inactivated seasonal influenza vaccine from the 2004-2005 season; the induction of NA titers was statistically less pronounced than the induction of HA titers. The demonstrated homologous and heterologous protective capacity of recombinant NA suggests that supplementing vaccine formulations with a standard amount of NA may offer increased protection against influenza virus infection.
Importance: Despite the existence of vaccine prophylaxis and antiviral therapeutics, the influenza virus continues to cause morbidity and mortality in the human population, emphasizing the continued need for research in the field. While the majority of influenza vaccine strategies target the viral hemagglutinin, the immunodominant antigen on the surface of the influenza virion, antibodies against the viral neuraminidase (NA) have been correlated with less severe disease and decreased viral shedding in humans. Nevertheless, the amount of NA is not standardized in current seasonal vaccines, and the exact breadth of NA-based protection is unknown. Greater insight into the cross-protective potential of influenza virus NA as a vaccine antigen may pave the way for the development of influenza vaccines of greater breadth and efficacy.
Copyright © 2015 Wohlbold et al.
Figures






References
-
- De Jong JC, Beyer WE, Palache AM, Rimmelzwaan GF, Osterhaus AD. 2000. Mismatch between the 1997/1998 influenza vaccine and the major epidemic A(H3N2) virus strain as the cause of an inadequate vaccine-induced antibody response to this strain in the elderly. J Med Virol 61:94–99. doi: 10.1002/(SICI)1096-9071(200005)61:1<94::AID-JMV15>3.0.CO;2-C. - DOI - PubMed
-
- Wrammert J, Smith K, Miller J, Langley WA, Kokko K, Larsen C, Zheng NY, Mays I, Garman L, Helms C, James J, Air GM, Capra JD, Ahmed R, Wilson PC. 2008. Rapid cloning of high-affinity human monoclonal antibodies against influenza virus. Nature 453:667–671. doi: 10.1038/nature06890. - DOI - PMC - PubMed
-
- Moody MA, Zhang R, Walter EB, Woods CW, Ginsburg GS, McClain MT, Denny TN, Chen X, Munshaw S, Marshall DJ, Whitesides JF, Drinker MS, Amos JD, Gurley TC, Eudailey JA, Foulger A, DeRosa KR, Parks R, Meyerhoff RR, Yu JS, Kozink DM, Barefoot BE, Ramsburg EA, Khurana S, Golding H, Vandergrift NA, Alam SM, Tomaras GD, Kepler TB, Kelsoe G, Liao HX, Haynes BF. 2011. H3N2 influenza infection elicits more cross-reactive and less clonally expanded anti-hemagglutinin antibodies than influenza vaccination. PLoS One 6:e25797. doi: 10.1371/journal.pone.0025797. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
