

The inhibitory effects of Geranium thunbergii on interferon-γ- and LPS-induced inflammatory responses are mediated by Nrf2 activation
- PMID: 25761198
- PMCID: PMC4380198
- DOI: 10.3892/ijmm.2015.2128
The inhibitory effects of Geranium thunbergii on interferon-γ- and LPS-induced inflammatory responses are mediated by Nrf2 activation
Abstract
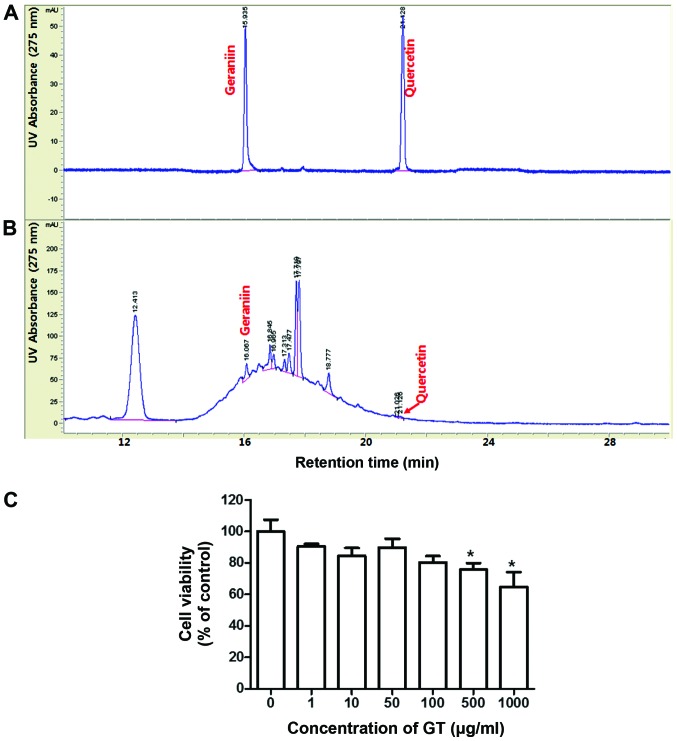
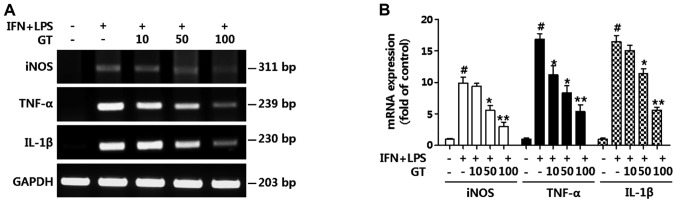
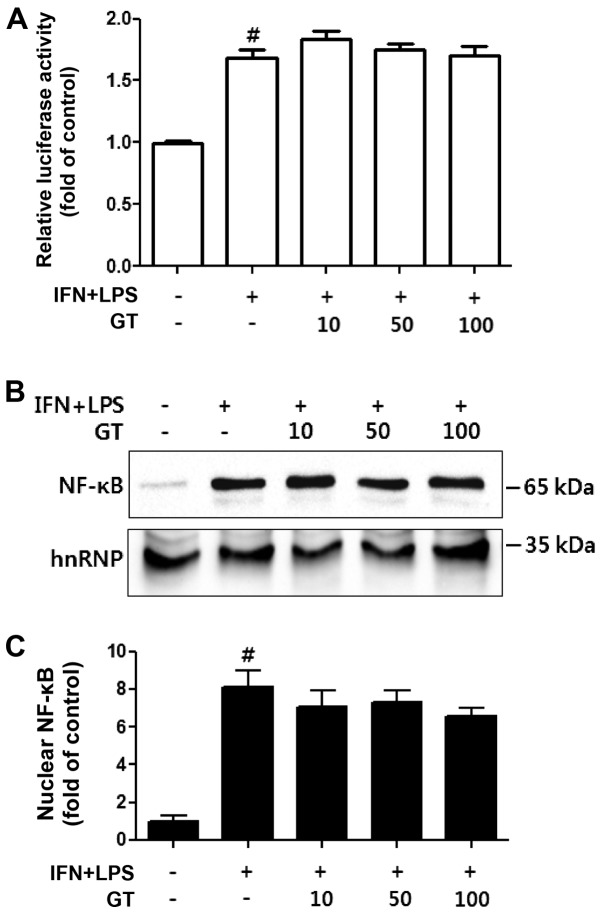
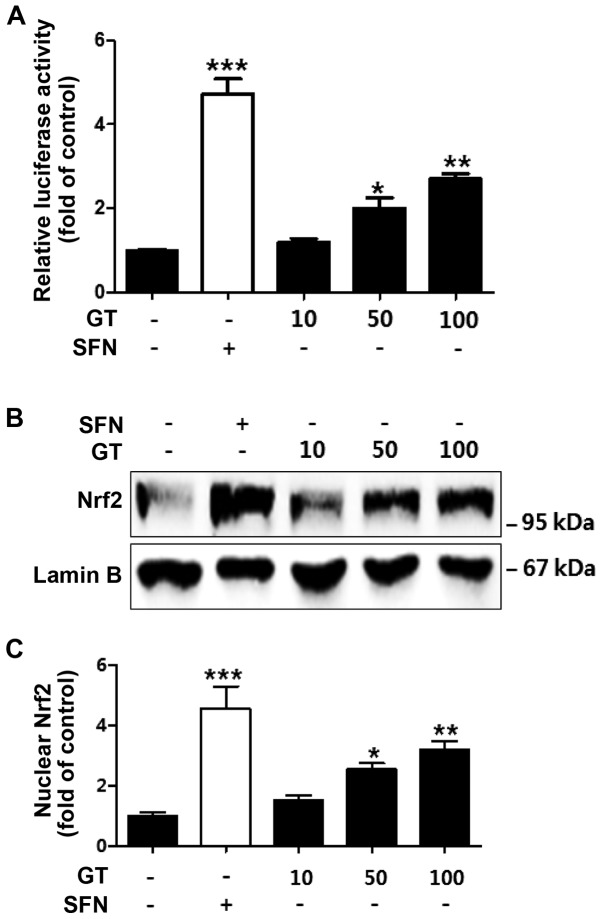
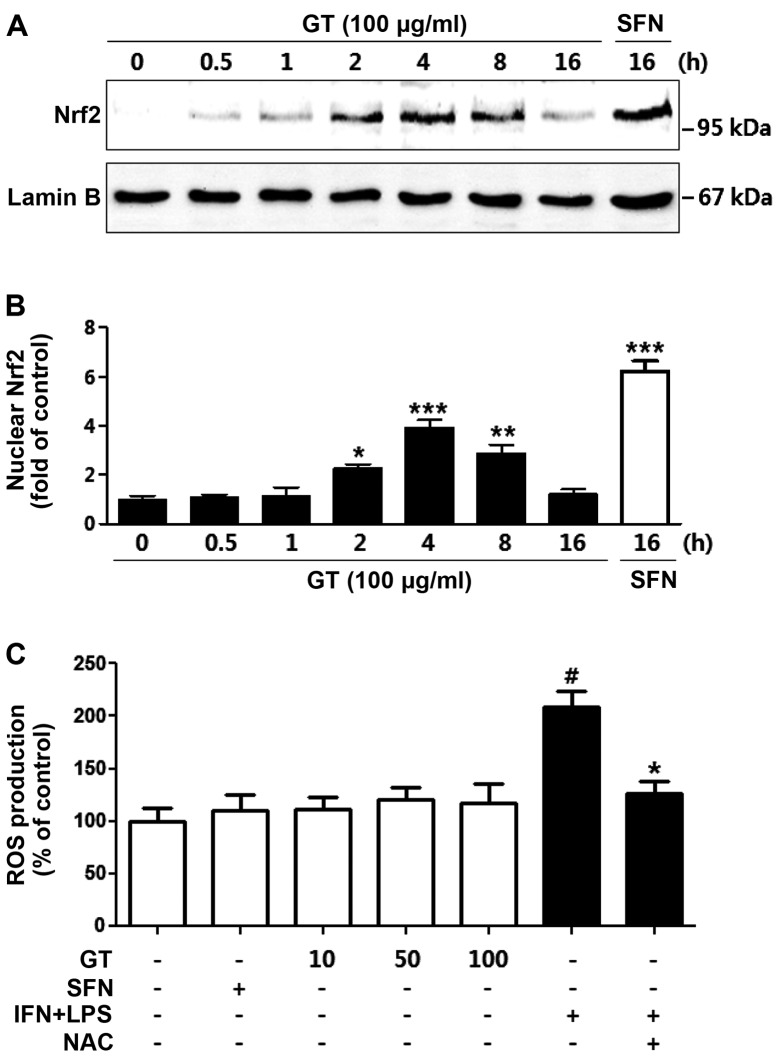
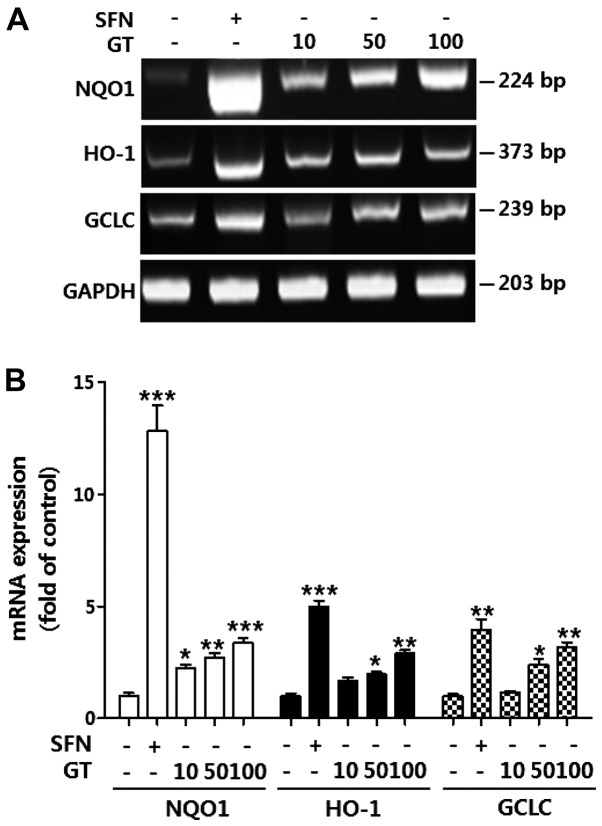
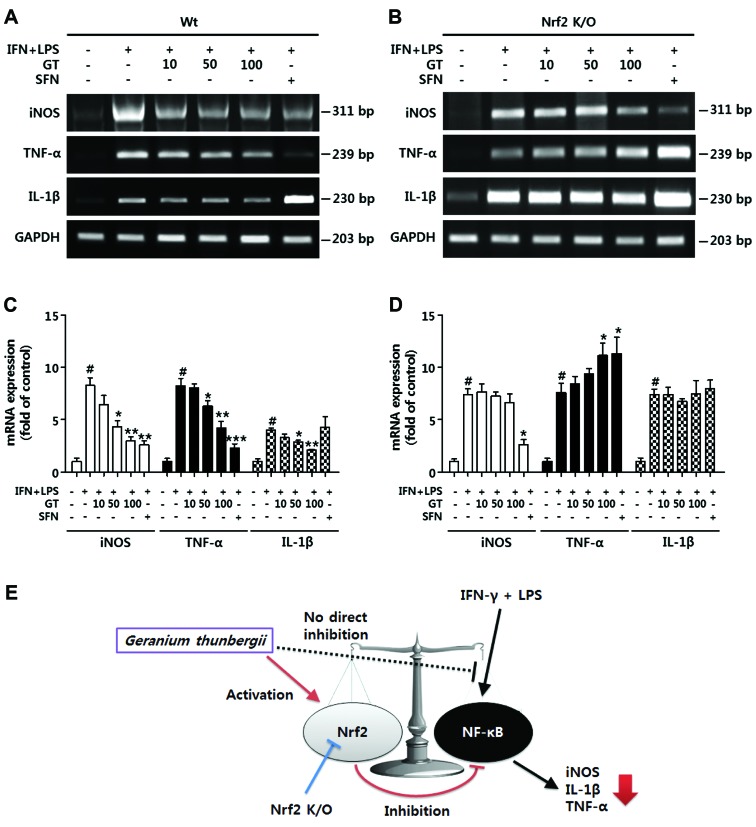
Geranium thunbergii Sieb. et Zucc. (GT; which belongs to the Geraniaceae family) has been used as a traditional medicine in East Asia for the treatment of inflammatory diseases, including arthritis and diarrhea. However, the underlying mechanisms of the anti-inflammatory effects of GT remain poorly understood. In the present study, we examined the mechanisms responsible for the anti-inflammatory activity of GT in macrophages. The results revealed that GT significantly inhibited the lipopolysaccharide (LPS)- and interferon-γ (IFN-γ)-induced expression of pro-inflammatory genes, such as inducible nitric oxide synthase, tumor necrosis factor-α and interleukin-1β, as shown by RT-PCR. However, the inhibitory effects of GT on LPS- and IFN-γ-induced inflammation were associated with an enhanced nuclear factor erythroid 2-related factor 2 (Nrf2) activity, but not with the suppression of nuclear factor (NF)-κB activity, as shown by western blot analysis. In addition, in bone marrow-derived macrophages (BMDM) isolated from Nrf2 knockout mice, GT did not exert any inhibitory effect on the LPS- and IFN-γ-induced inflammation. Taken together, our findings indicate that the anti-inflammatory effects of GT may be associated with the activation of Nrf2, an anti-inflammatory transcription factor.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
