

Cockayne syndrome group B (Csb) and group a (Csa) deficiencies predispose to hearing loss and cochlear hair cell degeneration in mice
- PMID: 25762674
- PMCID: PMC4355199
- DOI: 10.1523/JNEUROSCI.5063-14.2015
Cockayne syndrome group B (Csb) and group a (Csa) deficiencies predispose to hearing loss and cochlear hair cell degeneration in mice
Abstract
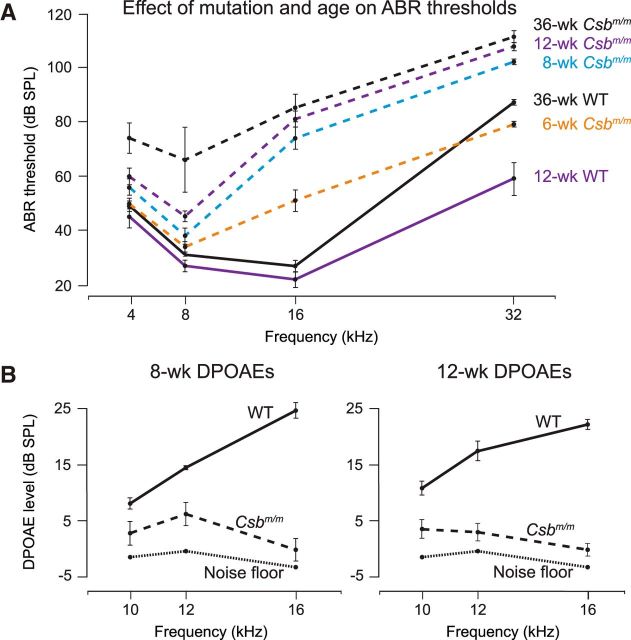
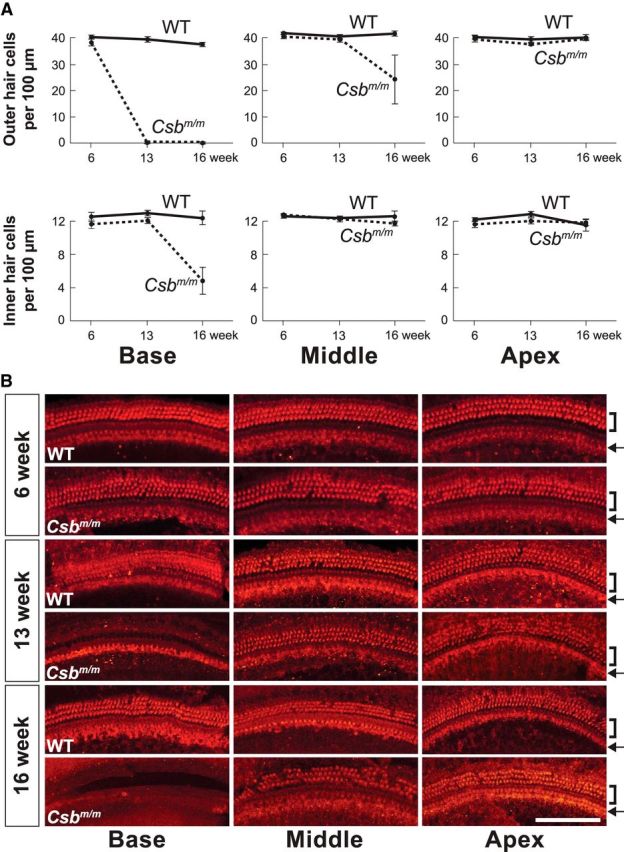
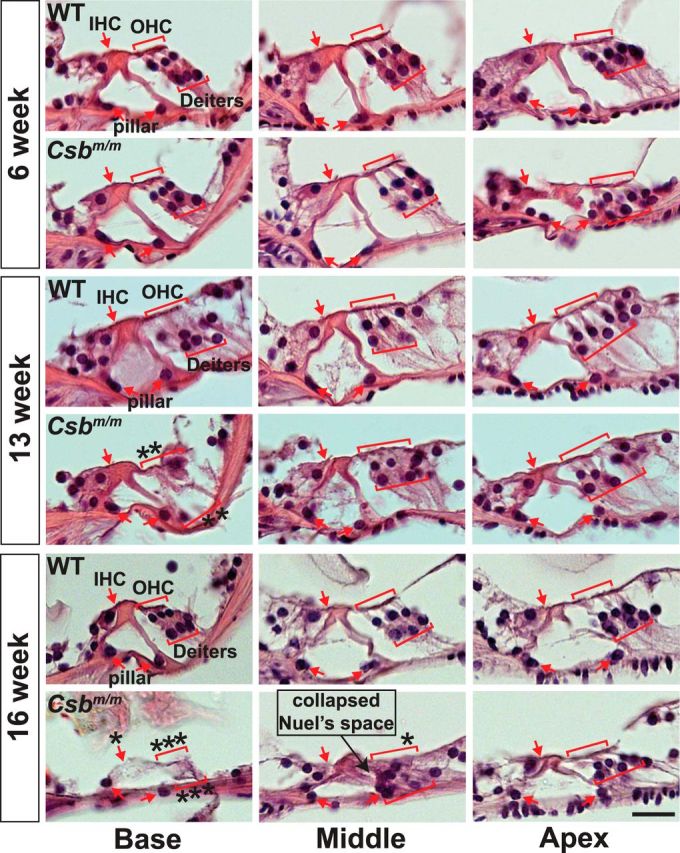
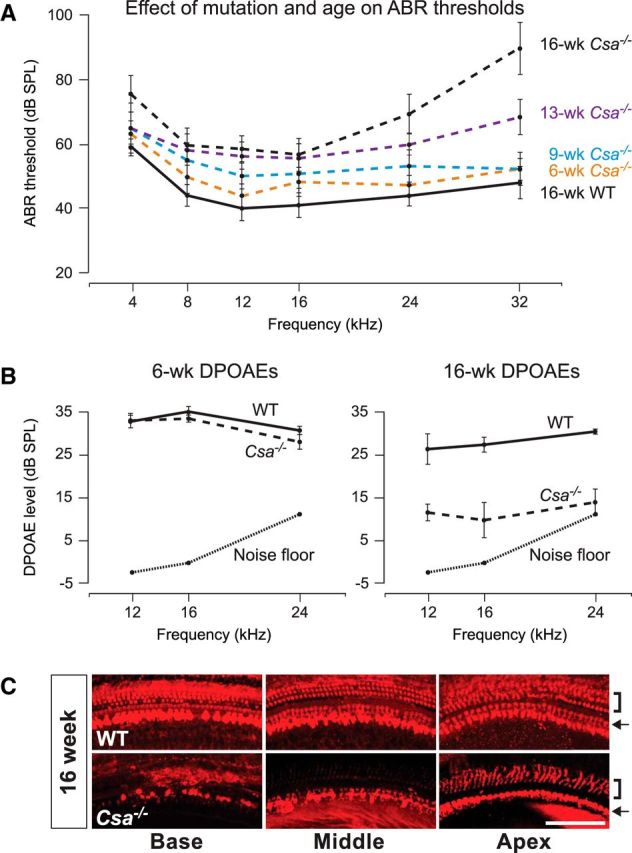
Sensory hair cells in the cochlea, like most neuronal populations that are postmitotic, terminally differentiated, and non-regenerating, depend on robust mechanisms of self-renewal for lifelong survival. We report that hair cell homeostasis requires a specific sub-branch of the DNA damage nucleotide excision repair pathway, termed transcription-coupled repair (TCR). Cockayne syndrome (CS), caused by defects in TCR, is a rare DNA repair disorder with a broad clinical spectrum that includes sensorineural hearing loss. We tested hearing and analyzed the cellular integrity of the organ of Corti in two mouse models of this disease with mutations in the Csb gene (CSB(m/m) mice) and Csa gene (Csa(-/-) mice), respectively. Csb(m/m) and Csa(-/-) mice manifested progressive hearing loss, as measured by an increase in auditory brainstem response thresholds. In contrast to wild-type mice, mutant mice showed reduced or absent otoacoustic emissions, suggesting cochlear outer hair cell impairment. Hearing loss in Csb(m/m) and Csa(-/-) mice correlated with progressive hair cell loss in the base of the organ of Corti, starting between 6 and 13 weeks of age, which increased by 16 weeks of age in a basal-to-apical gradient, with outer hair cells more severely affected than inner hair cells. Our data indicate that the hearing loss observed in CS patients is reproduced in mouse models of this disease. We hypothesize that accumulating DNA damage, secondary to the loss of TCR, contributes to susceptibility to hearing loss.
Keywords: Cockayne syndrome; DNA damage; DNA repair; csb/csa; hair cell; hearing loss.
Copyright © 2015 the authors 0270-6474/15/354280-07$15.00/0.
Figures
References
-
- Berg RJ, Rebel H, van der Horst GT, van Kranen HJ, Mullenders LH, van Vloten WA, de Gruijl FR. Impact of global genome repair versus transcription-coupled repair on ultraviolet carcinogenesis in hairless mice. Cancer Res. 2000;60:2858–2863. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases