

Murine Gammaherpesvirus 68 ORF48 Is an RTA-Responsive Gene Product and Functions in both Viral Lytic Replication and Latency during In Vivo Infection
- PMID: 25762743
- PMCID: PMC4442421
- DOI: 10.1128/JVI.00406-15
Murine Gammaherpesvirus 68 ORF48 Is an RTA-Responsive Gene Product and Functions in both Viral Lytic Replication and Latency during In Vivo Infection
Abstract
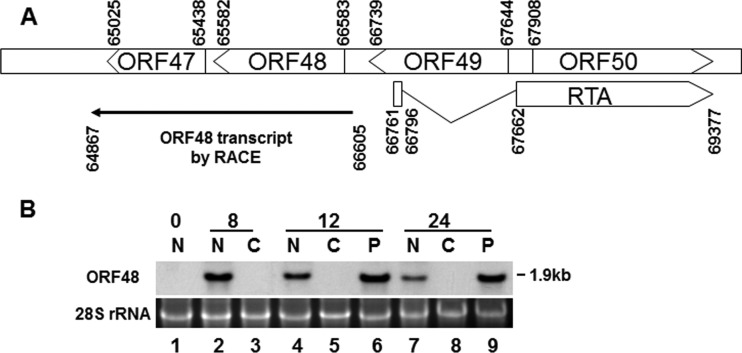
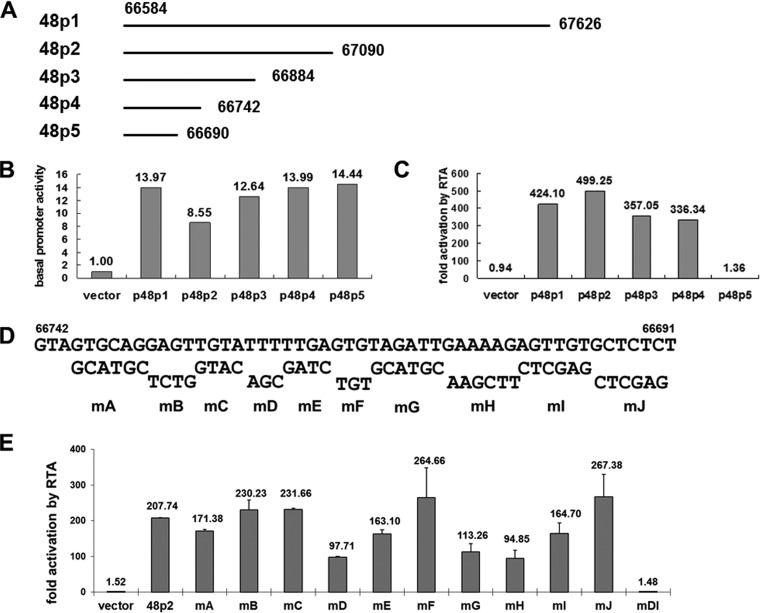
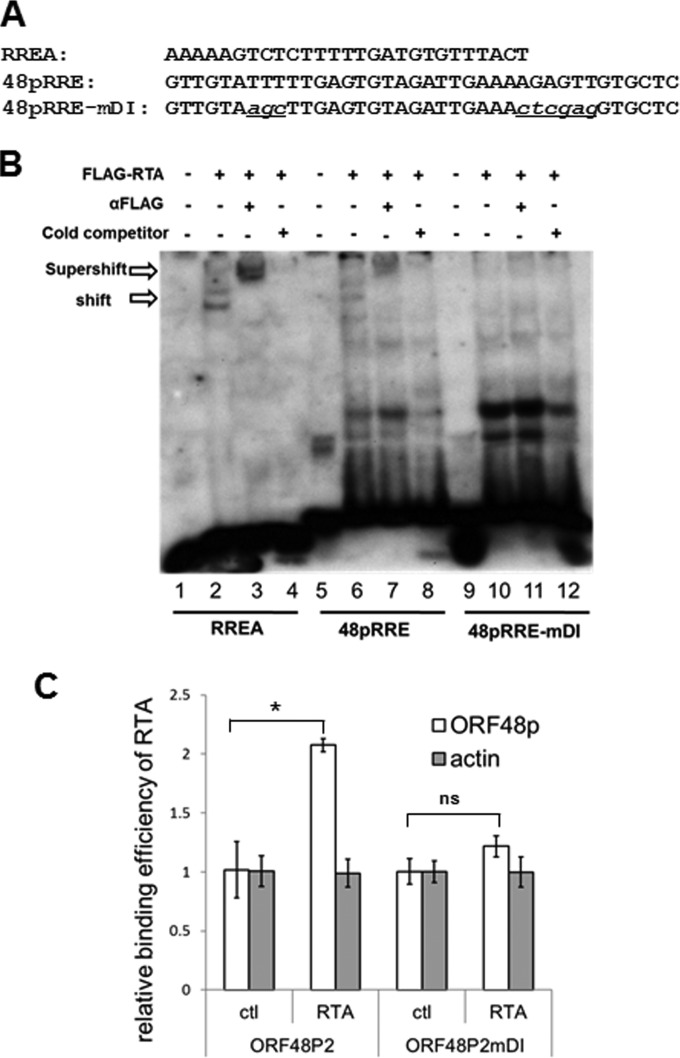
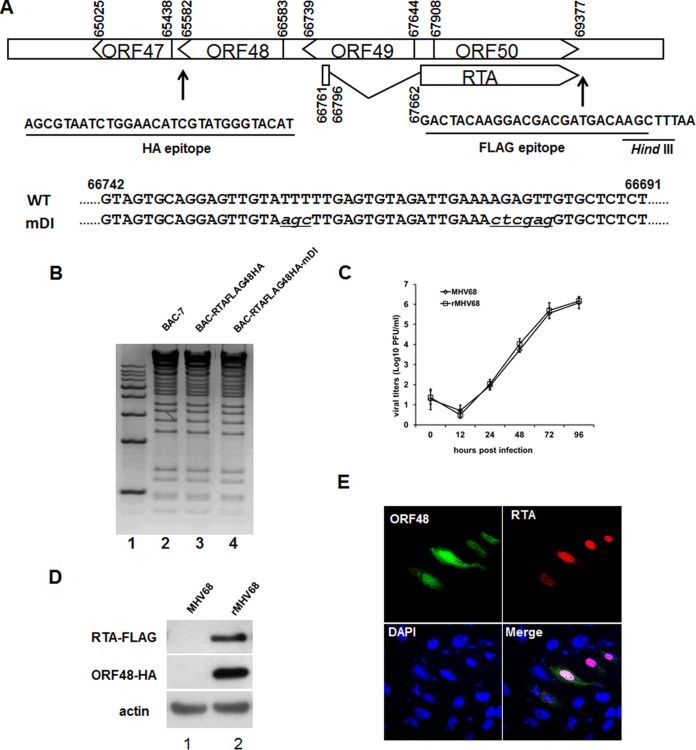
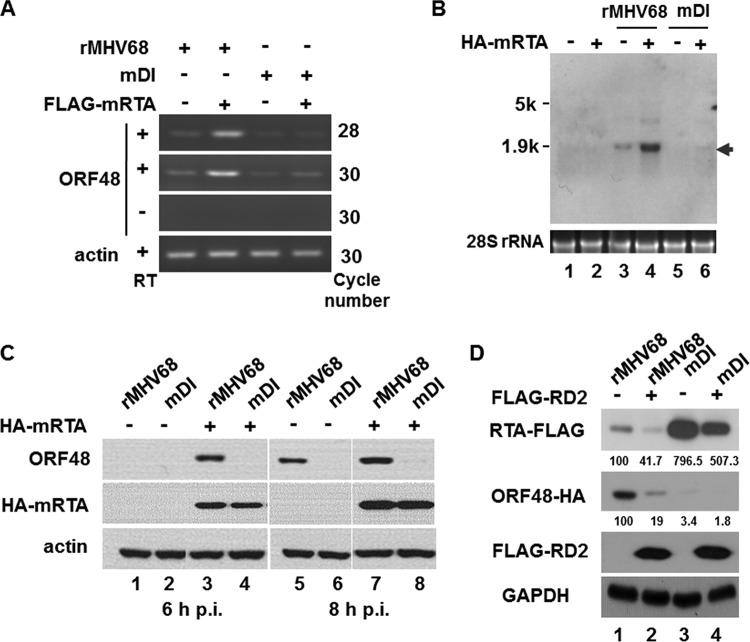
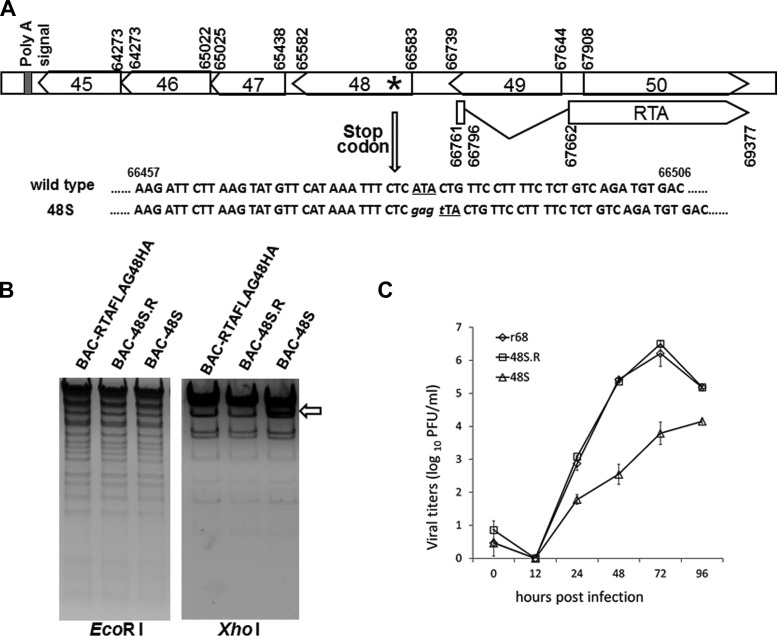
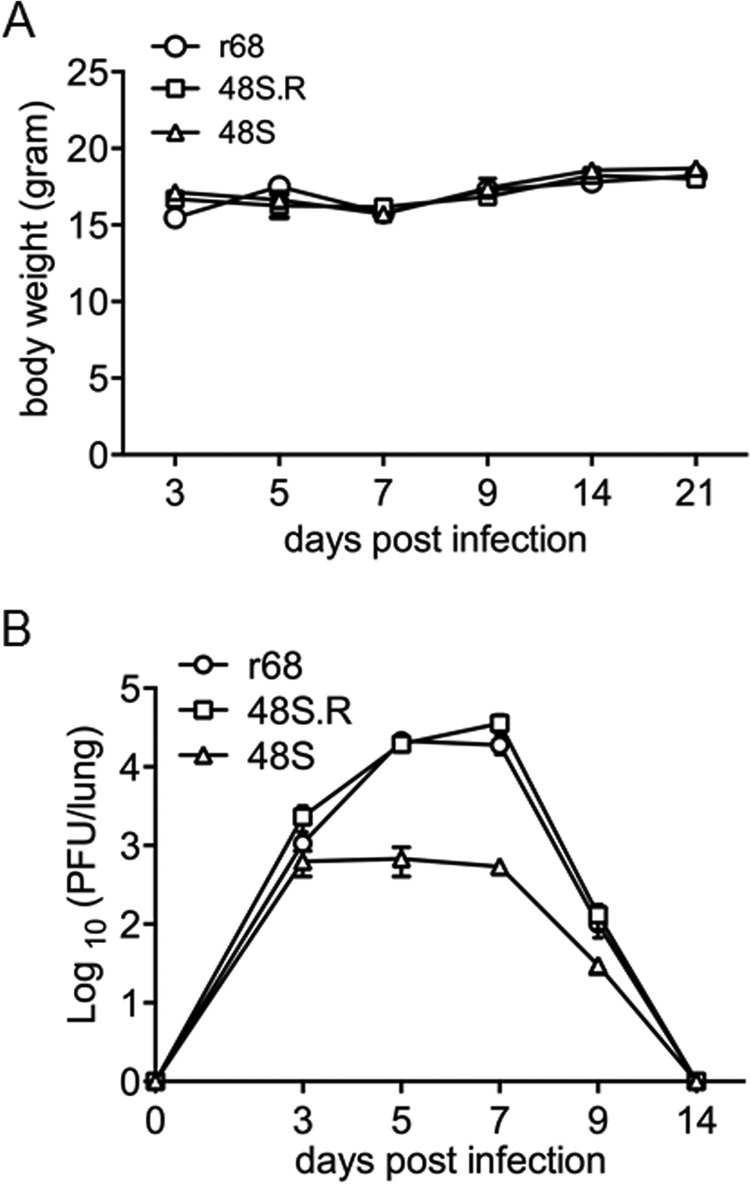
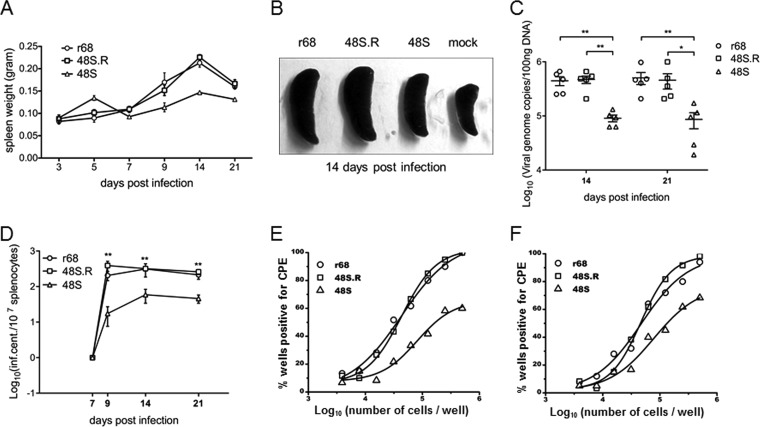
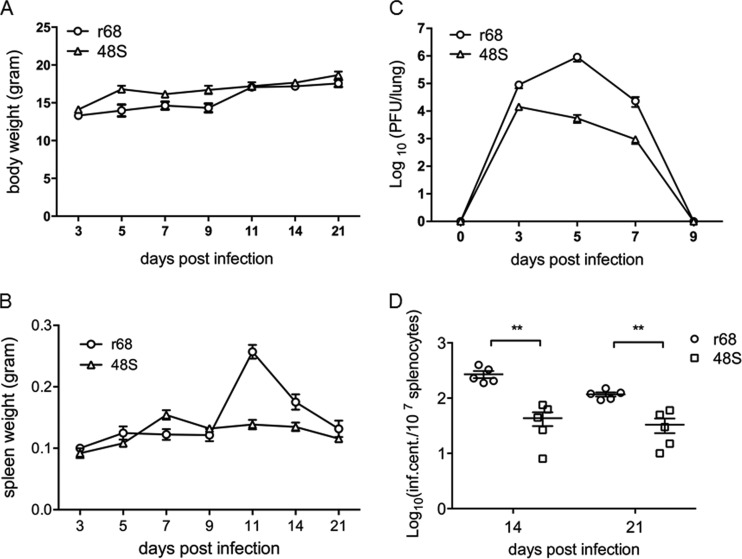
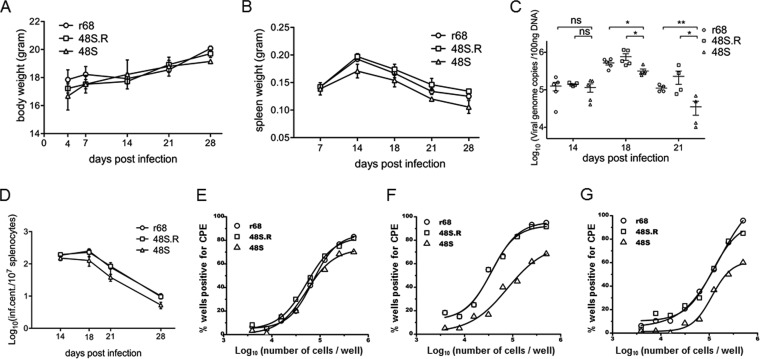
Replication and transcription activator (RTA) of gammaherpesvirus is an immediate early gene product and regulates the expression of many downstream viral lytic genes. ORF48 is also conserved among gammaherpesviruses; however, its expression regulation and function remained largely unknown. In this study, we characterized the transcription unit of ORF48 from murine gammaherpesvirus 68 (MHV-68) and analyzed its transcriptional regulation. We showed that RTA activates the ORF48 promoter via an RTA-responsive element (48pRRE). RTA binds to 48pRRE directly in vitro and also associates with ORF48 promoter in vivo. Mutagenesis of 48pRRE in the context of the viral genome demonstrated that the expression of ORF48 is activated by RTA through 48pRRE during de novo infection. Through site-specific mutagenesis, we generated an ORF48-null virus and examined the function of ORF48 in vitro and in vivo. The ORF48-null mutation remarkably reduced the viral replication efficiency in cell culture. Moreover, through intranasal or intraperitoneal infection of laboratory mice, we showed that ORF48 is important for viral lytic replication in the lung and establishment of latency in the spleen, as well as viral reactivation from latency. Collectively, our study identified ORF48 as an RTA-responsive gene and showed that ORF48 is important for MHV-68 replication both in vitro and in vivo.
Importance: The replication and transcription activator (RTA), conserved among gammaherpesviruses, serves as a molecular switch for the virus life cycle. It works as a transcriptional regulator to activate the expression of many viral lytic genes. However, only a limited number of such downstream genes have been uncovered for MHV-68. In this study, we identified ORF48 as an RTA-responsive gene of MHV-68 and mapped the cis element involved. By constructing a mutant virus that is deficient in ORF48 expression and through infection of laboratory mice, we showed that ORF48 plays important roles in different stages of viral infection in vivo. Our study provides insights into the transcriptional regulation and protein function of MHV-68, a desired model for studying gammaherpesviruses.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
