

Coexpression of TIGIT and FCRL3 identifies Helios+ human memory regulatory T cells
- PMID: 25762785
- PMCID: PMC4610024
- DOI: 10.4049/jimmunol.1401803
Coexpression of TIGIT and FCRL3 identifies Helios+ human memory regulatory T cells
Abstract
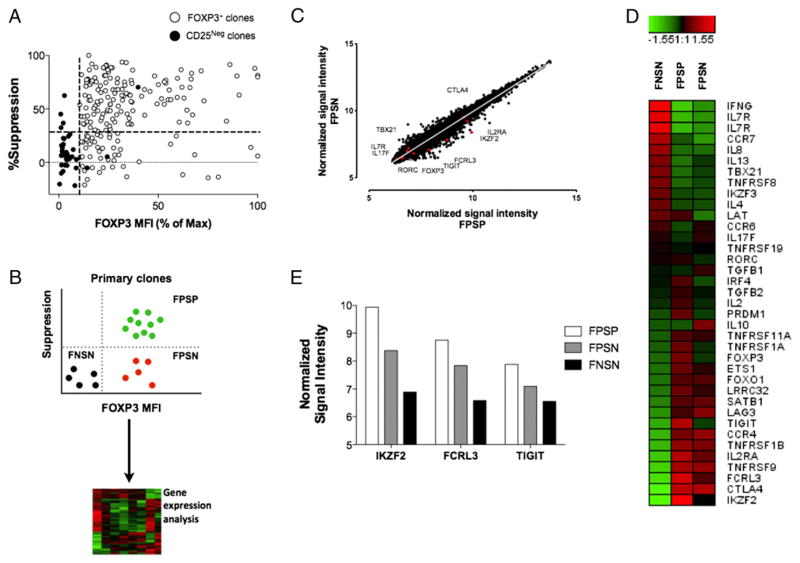
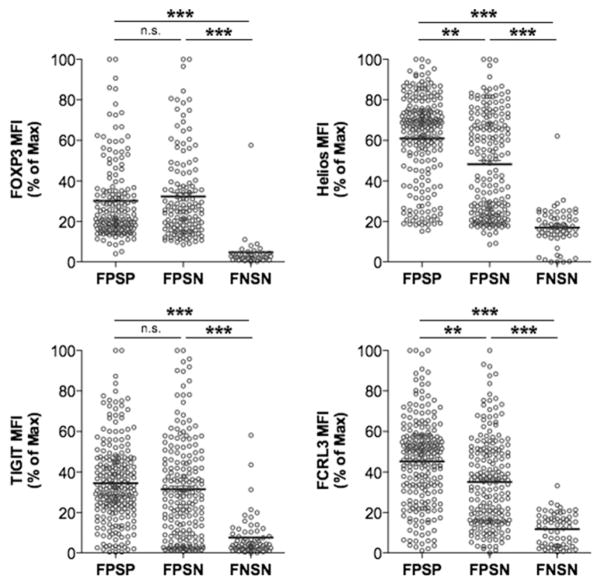
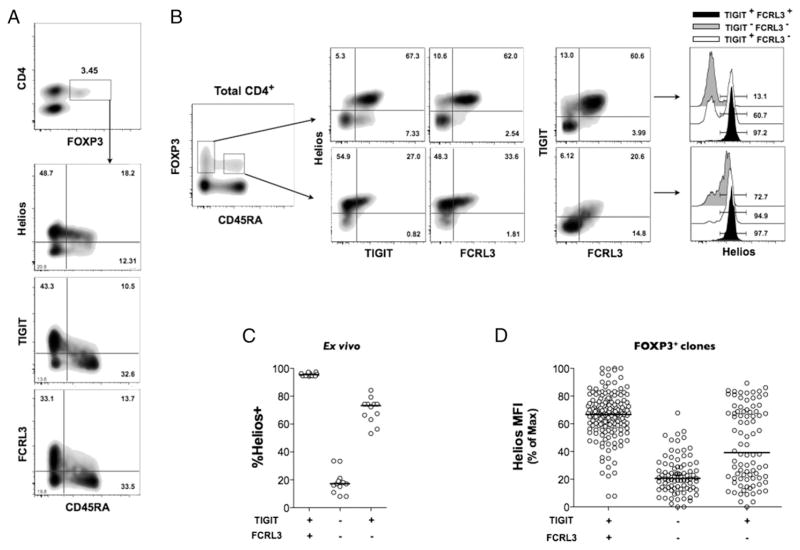
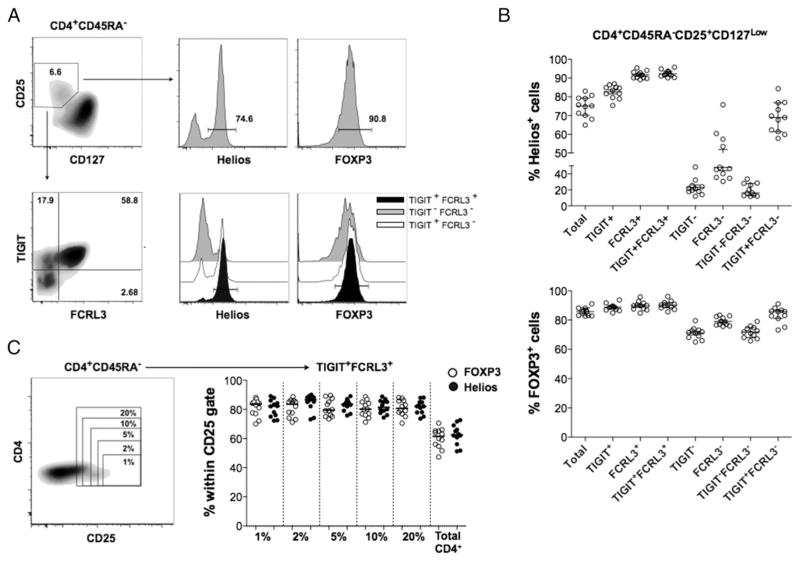
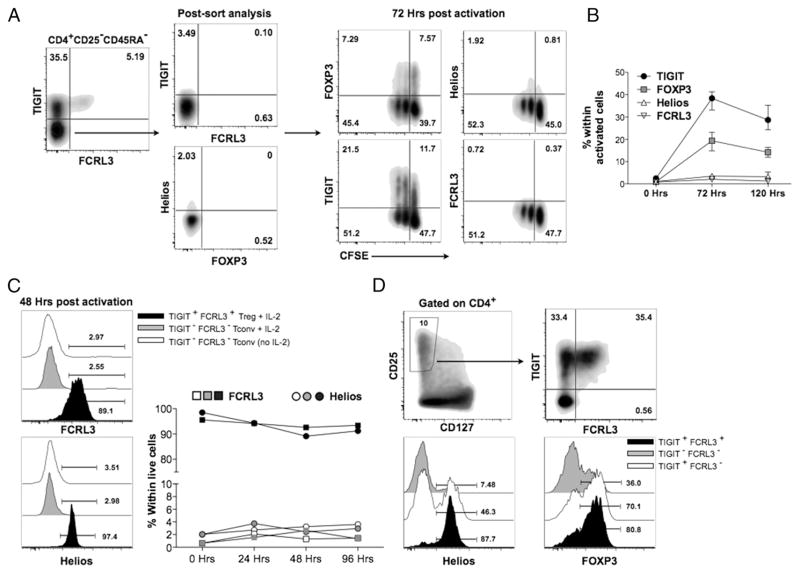
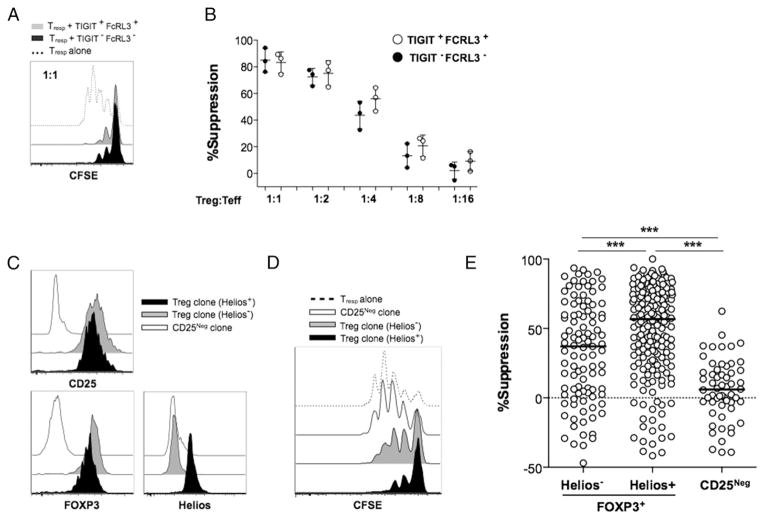
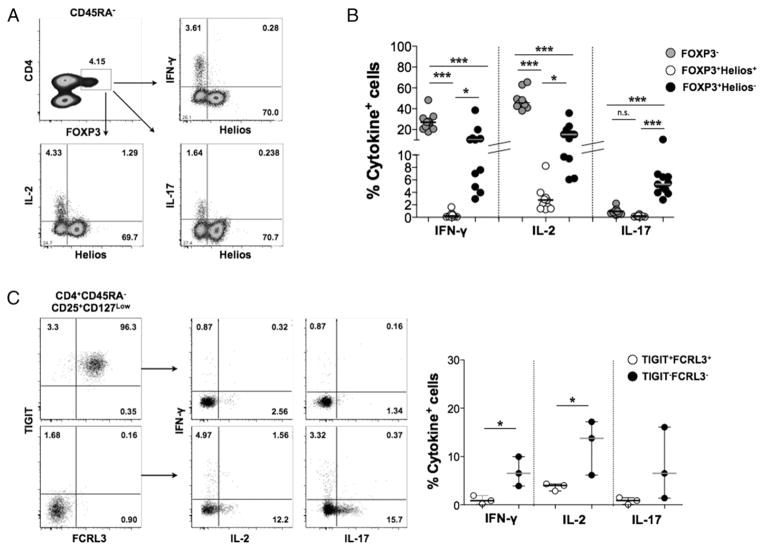
Two distinct subsets of CD4(+)Foxp3(+) regulatory T (Treg) cells have been described based on the differential expression of Helios, a transcription factor of the Ikaros family. Efforts to understand the origin and biological roles of these Treg populations in regulating immune responses have, however, been hindered by the lack of reliable surface markers to distinguish and isolate them for subsequent functional studies. Using a single-cell cloning strategy coupled with microarray analysis of different Treg functional subsets in humans, we identify the mRNA and protein expression of TIGIT and FCRL3 as a novel surface marker combination that distinguishes Helios(+)FOXP3(+) from Helios(-)FOXP3(+) memory cells. Unlike conventional markers that are modulated on conventional T cells upon activation, we show that the TIGIT/FCRL3 combination allows reliable identification of Helios(+) Treg cells even in highly activated conditions in vitro as well as in PBMCs of autoimmune patients. We also demonstrate that the Helios(-)FOXP3(+) Treg subpopulation harbors a larger proportion of nonsuppressive clones compared with the Helios(+)FOXP3(+) cell subset, which is highly enriched for suppressive clones. Moreover, we find that Helios(-) cells are exclusively responsible for the productions of the inflammatory cytokines IFN-γ, IL-2, and IL-17 in FOXP3(+) cells ex vivo, highlighting important functional differences between Helios(+) and Helios(-) Treg cells. Thus, we identify novel surface markers for the consistent identification and isolation of Helios(+) and Helios(-) memory Treg cells in health and disease, and we further reveal functional differences between these two populations. These new markers should facilitate further elucidation of the functional roles of Helios-based Treg heterogeneity.
Copyright © 2015 by The American Association of Immunologists, Inc.
Conflict of interest statement
The authors have no financial conflicts of interest.
Figures
References
-
- Sakaguchi S, Miyara M, Costantino CM, Hafler DA. FOXP3+ regulatory T cells in the human immune system. Nat Rev Immunol. 2010;10:490–500. - PubMed
-
- d’Hennezel E, Bin Dhuban K, Torgerson T, Piccirillo CA. The immunogenetics of immune dysregulation, polyendocrinopathy, enteropathy, X linked (IPEX) syndrome. J Med Genet. 2012;49:291–302. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
