

Accumulation of squalene in a microalga Chlamydomonas reinhardtii by genetic modification of squalene synthase and squalene epoxidase genes
- PMID: 25764133
- PMCID: PMC4357444
- DOI: 10.1371/journal.pone.0120446
Accumulation of squalene in a microalga Chlamydomonas reinhardtii by genetic modification of squalene synthase and squalene epoxidase genes
Abstract
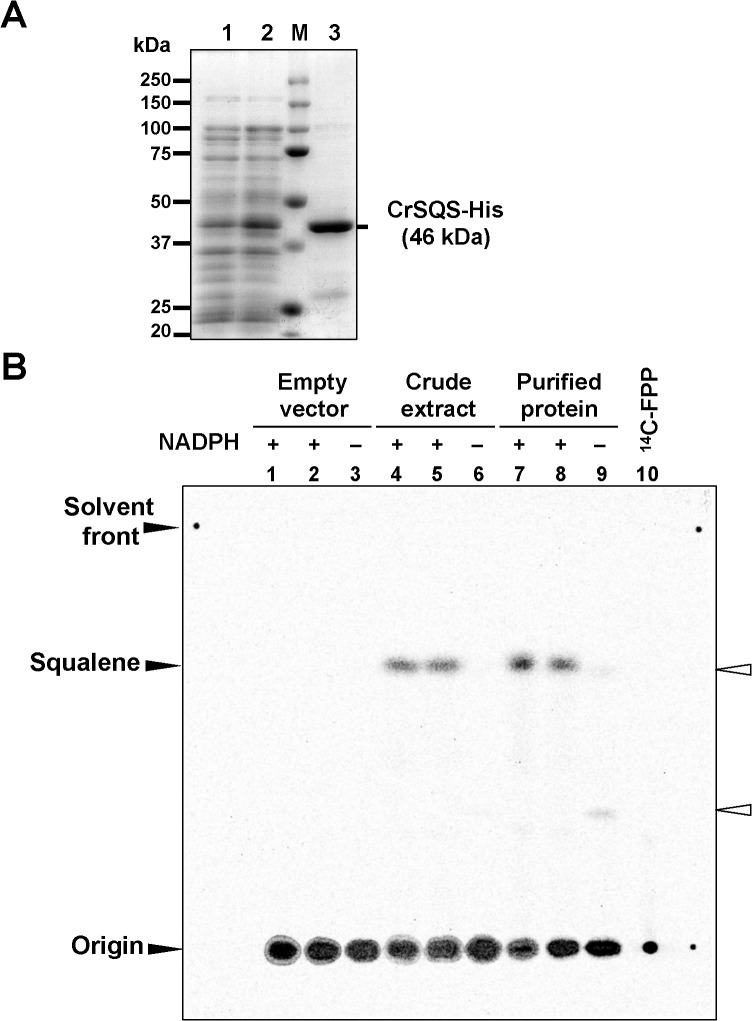
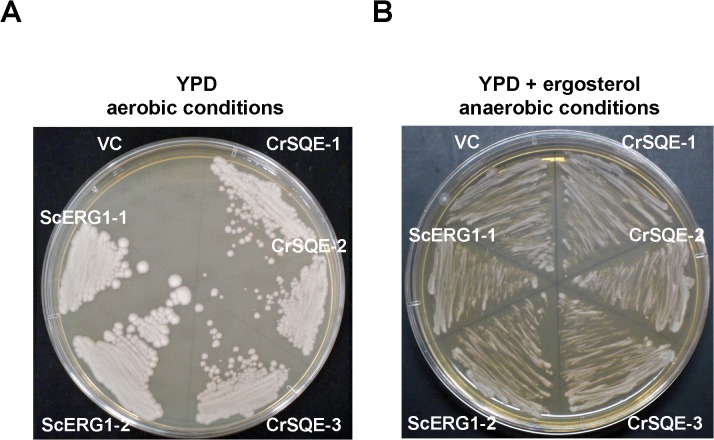
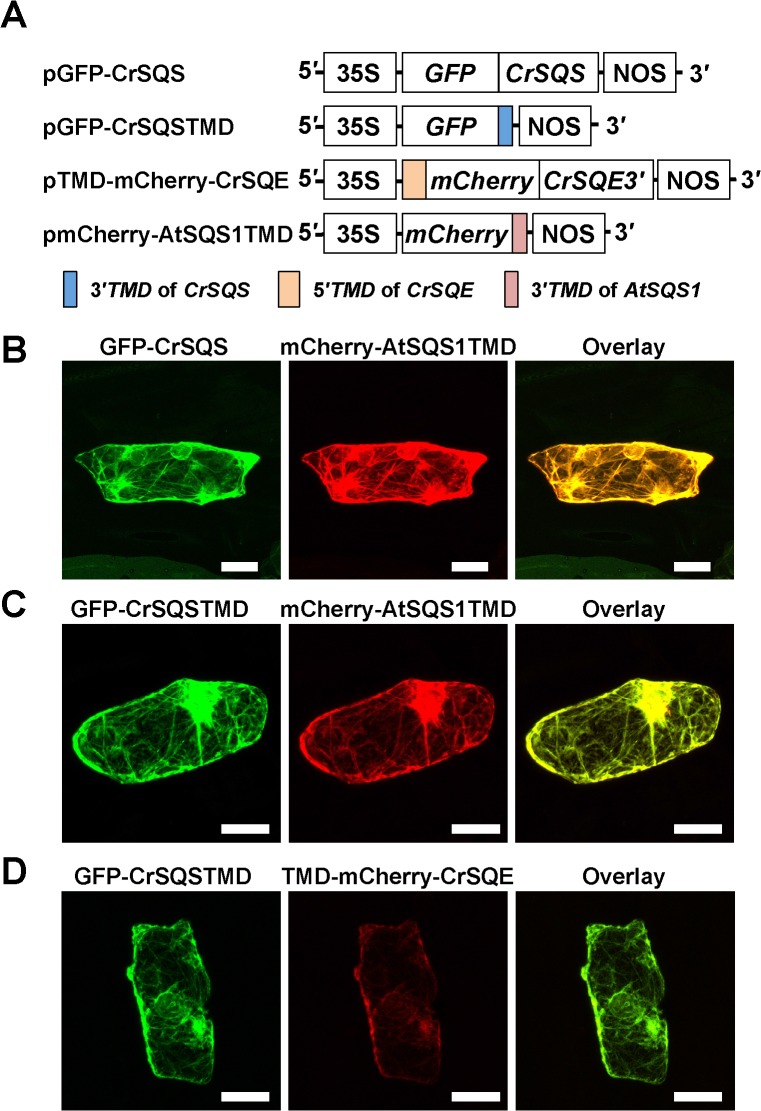
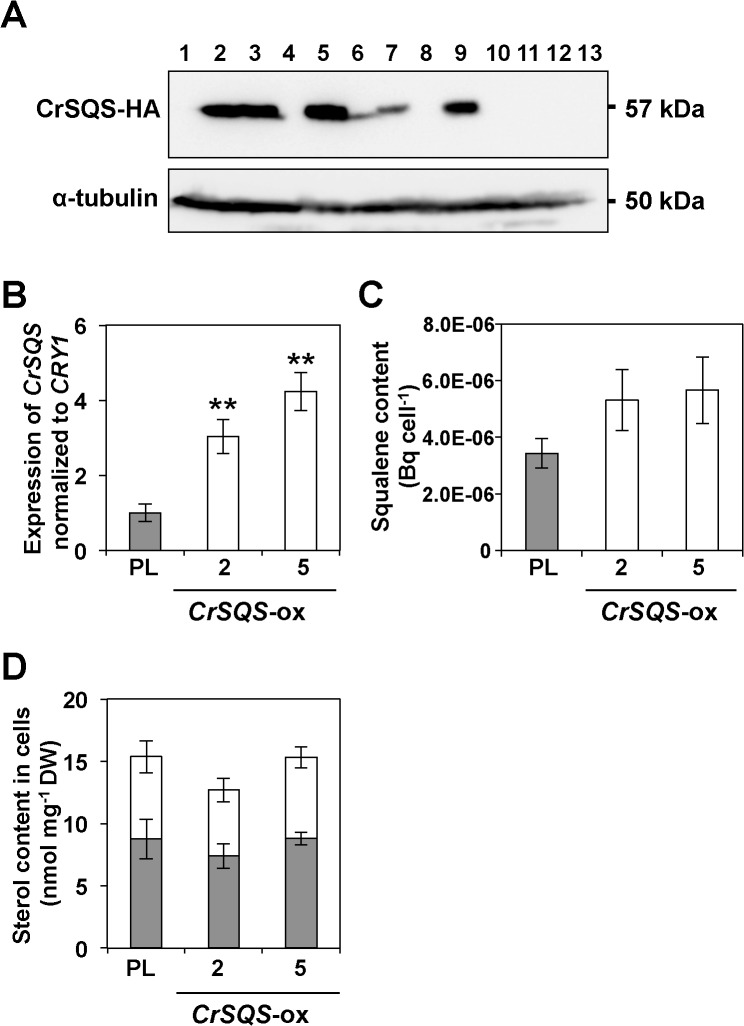
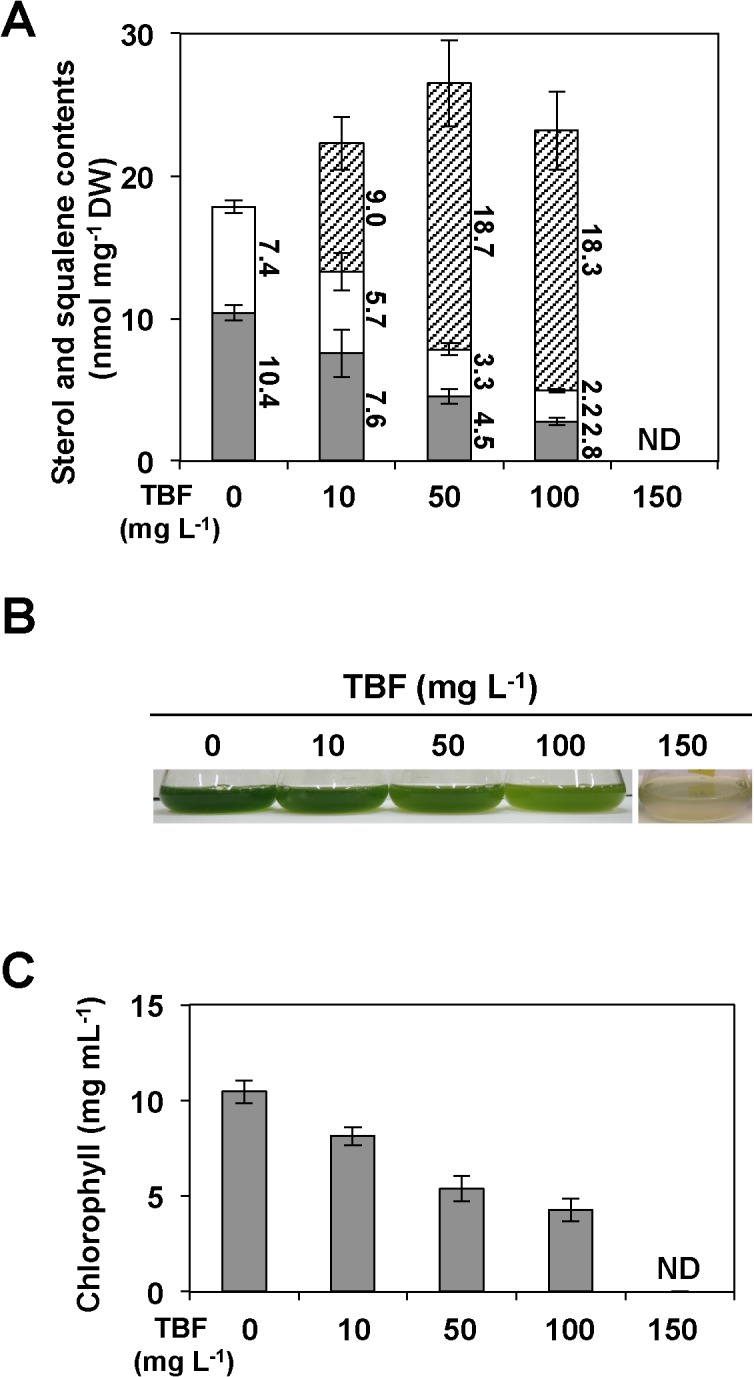
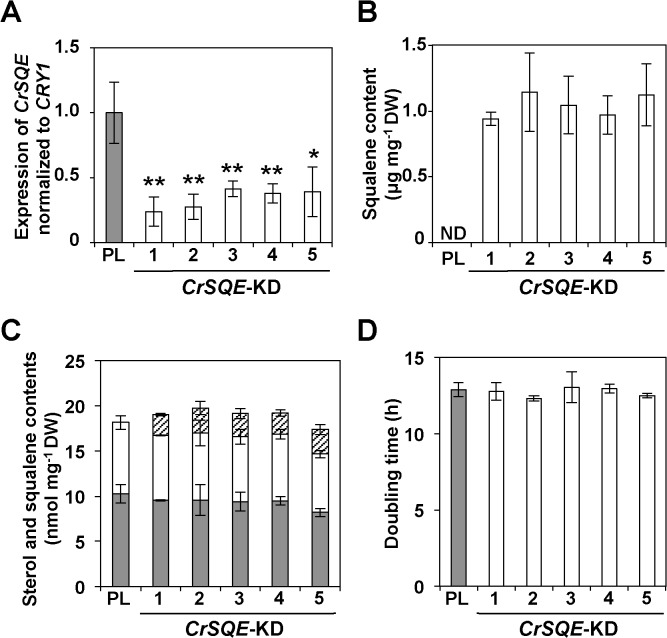
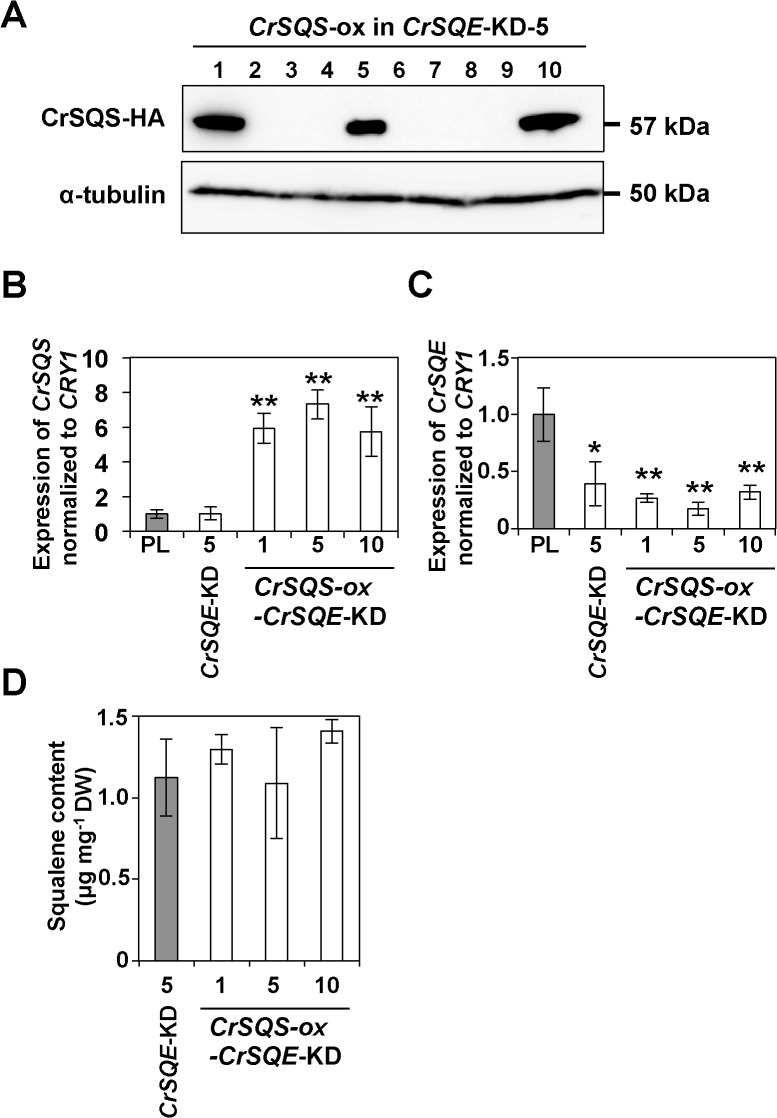
Several microalgae accumulate high levels of squalene, and as such provide a potentially valuable source of this useful compound. However, the molecular mechanism of squalene biosynthesis in microalgae is still largely unknown. We obtained the sequences of two enzymes involved in squalene synthesis and metabolism, squalene synthase (CrSQS) and squalene epoxidase (CrSQE), from the model green alga Chlamydomonas reinhardtii. CrSQS was functionally characterized by expression in Escherichia coli and CrSQE by complementation of a budding yeast erg1 mutant. Transient expression of CrSQS and CrSQE fused with fluorescent proteins in onion epidermal tissue suggested that both proteins were co-localized in the endoplasmic reticulum. CrSQS-overexpression increased the rate of conversion of 14C-labeled farnesylpyrophosphate into squalene but did not lead to over-accumulation of squalene. Addition of terbinafine caused the accumulation of squalene and suppression of cell survival. On the other hand, in CrSQE-knockdown lines, the expression level of CrSQE was reduced by 59-76% of that in wild-type cells, and significant levels of squalene (0.9-1.1 μg mg-1 cell dry weight) accumulated without any growth inhibition. In co-transformation lines with CrSQS-overexpression and CrSQE-knockdown, the level of squalene was not increased significantly compared with that in solitary CrSQE-knockdown lines. These results indicated that partial knockdown of CrSQE is an effective strategy to increase squalene production in C. reinhardtii cells.
Conflict of interest statement
Figures
Similar articles
-
Inhibition of squalene synthase and squalene epoxidase in tobacco cells triggers an up-regulation of 3-hydroxy-3-methylglutaryl coenzyme a reductase.Plant Physiol. 2002 Sep;130(1):334-46. doi: 10.1104/pp.004655. Plant Physiol. 2002. PMID: 12226513 Free PMC article.
-
Squalene epoxidase as a target for manipulation of squalene levels in the yeast Saccharomyces cerevisiae.FEMS Yeast Res. 2014 Mar;14(2):310-23. doi: 10.1111/1567-1364.12107. Epub 2013 Oct 30. FEMS Yeast Res. 2014. PMID: 24119181
-
Dual localization of squalene epoxidase, Erg1p, in yeast reflects a relationship between the endoplasmic reticulum and lipid particles.Mol Biol Cell. 1998 Feb;9(2):375-86. doi: 10.1091/mbc.9.2.375. Mol Biol Cell. 1998. PMID: 9450962 Free PMC article.
-
Terbinafine: mode of action and properties of the squalene epoxidase inhibition.Br J Dermatol. 1992 Feb;126 Suppl 39:2-7. doi: 10.1111/j.1365-2133.1992.tb00001.x. Br J Dermatol. 1992. PMID: 1543672 Review.
-
Inhibitors of squalene biosynthesis and metabolism.Nat Prod Rep. 1994 Jun;11(3):279-302. doi: 10.1039/np9941100279. Nat Prod Rep. 1994. PMID: 15200015 Review. No abstract available.
Cited by
-
Bioprocess conditions and regulation factors to optimize squalene production in thraustochytrids.World J Microbiol Biotechnol. 2023 Jul 14;39(9):251. doi: 10.1007/s11274-023-03689-y. World J Microbiol Biotechnol. 2023. PMID: 37442840 Review.
-
Exploiting microalgal diversity for sterol production.Front Plant Sci. 2025 Jun 30;16:1616863. doi: 10.3389/fpls.2025.1616863. eCollection 2025. Front Plant Sci. 2025. PMID: 40661763 Free PMC article. Review.
-
Microbial Squalene: A Sustainable Alternative for the Cosmetics and Pharmaceutical Industry - A Review.Eng Life Sci. 2024 Aug 27;24(10):e202400003. doi: 10.1002/elsc.202400003. eCollection 2024 Oct. Eng Life Sci. 2024. PMID: 39391272 Free PMC article. Review.
-
Functional characterization of squalene synthase and squalene epoxidase in Taraxacum koksaghyz.Plant Direct. 2018 Jun 13;2(6):e00063. doi: 10.1002/pld3.63. eCollection 2018 Jun. Plant Direct. 2018. PMID: 31245726 Free PMC article.
-
In Metabolic Engineering of Eukaryotic Microalgae: Potential and Challenges Come with Great Diversity.Front Microbiol. 2015 Dec 15;6:1376. doi: 10.3389/fmicb.2015.01376. eCollection 2015. Front Microbiol. 2015. PMID: 26696985 Free PMC article. Review.
References
-
- Spanova M, Daum G. Squalene—biochemistry, molecular biology, process biotechnology, and applications. Eur J Lipid Sci Technol. 2011;113: 1299–1320.
-
- Bhattacharjee P, Shukla VB, Singhal RS, Kulkarni PR. Studies on fermentative production of squalene. World J Microbiol Biotechnol. 2001;17: 811–816.
-
- Jiang Y, Fan KW, Wong RDY, Chen F. Fatty acid composition and squalene content of the marine microalga Schizochytrium mangrovei . J Agric Food Chem. 2004;52: 1196–1200. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
