

Activation and reactivation of the RNA polymerase II trigger loop for intrinsic RNA cleavage and catalysis
- PMID: 25764335
- PMCID: PMC4574878
- DOI: 10.4161/trns.28869
Activation and reactivation of the RNA polymerase II trigger loop for intrinsic RNA cleavage and catalysis
Abstract
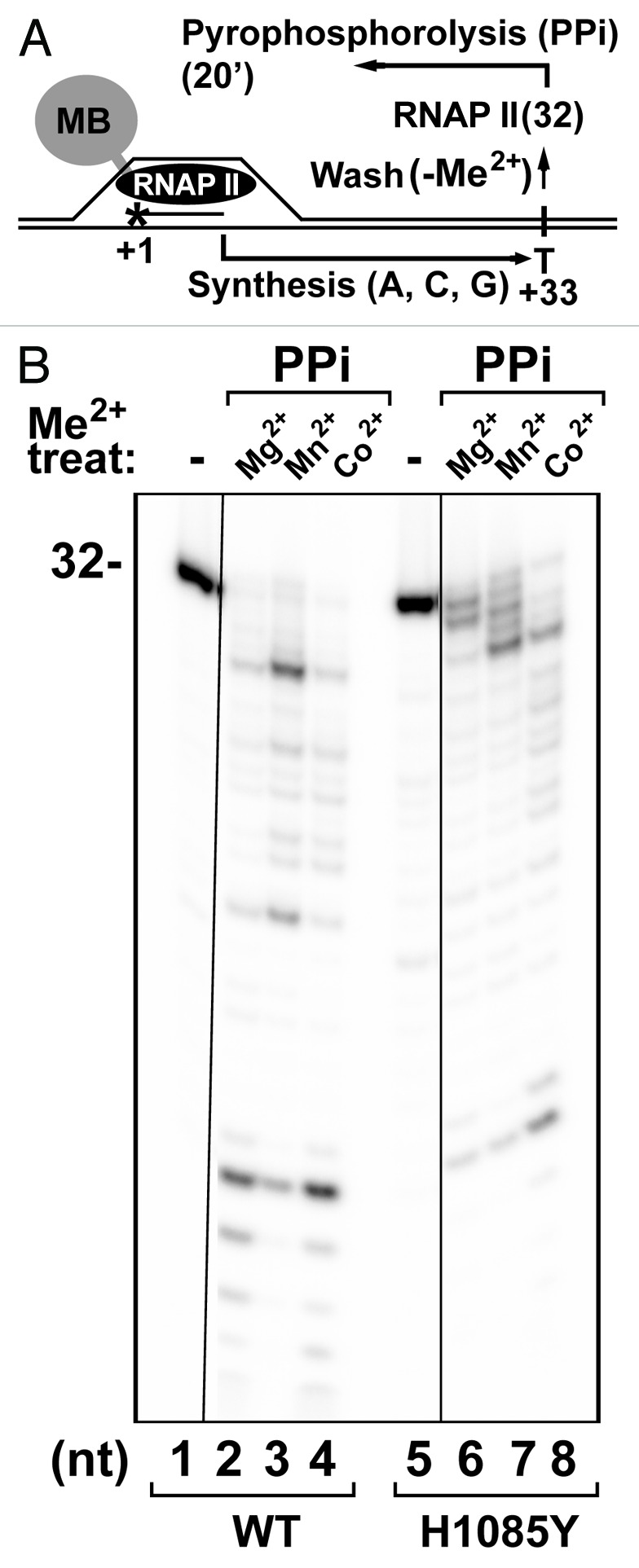
In addition to RNA synthesis, multisubunit RNA polymerases (msRNAPs) support enzymatic reactions such as intrinsic transcript cleavage. msRNAP active sites from different species appear to exhibit differential intrinsic transcript cleavage efficiency and have likely evolved to allow fine-tuning of the transcription process. Here we show that a single amino-acid substitution in the trigger loop (TL) of Saccharomyces RNAP II, Rpb1 H1085Y, engenders a gain of intrinsic cleavage activity where the substituted tyrosine appears to participate in acid-base chemistry at alkaline pH for both intrinsic cleavage and nucleotidyl transfer. We extensively characterize this TL substitution for each of these reactions by examining the responses RNAP II enzymes to catalytic metals, altered pH, and factor inputs. We demonstrate that TFIIF stimulation of the first phosphodiester bond formation by RNAP II requires wild type TL function and that H1085Y substitution within the TL compromises or alters RNAP II responsiveness to both TFIIB and TFIIF. Finally, Mn(2+) stimulation of H1085Y RNAP II reveals possible allosteric effects of TFIIB on the active center and cooperation between TFIIB and TFIIF.
Keywords: RNA synthesis; gene expression; general transcription factors; intrinsic cleavage; trigger loop.
Figures








References
-
- Whitehall SK, Bardeleben C, Kassavetis GA. Hydrolytic cleavage of nascent RNA in RNA polymerase III ternary transcription complexes. J Biol Chem. 1994;269:2299–306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases