

Autophagy Controls Acquisition of Aging Features in Macrophages
- PMID: 25764971
- PMCID: PMC4386145
- DOI: 10.1159/000370112
Autophagy Controls Acquisition of Aging Features in Macrophages
Abstract
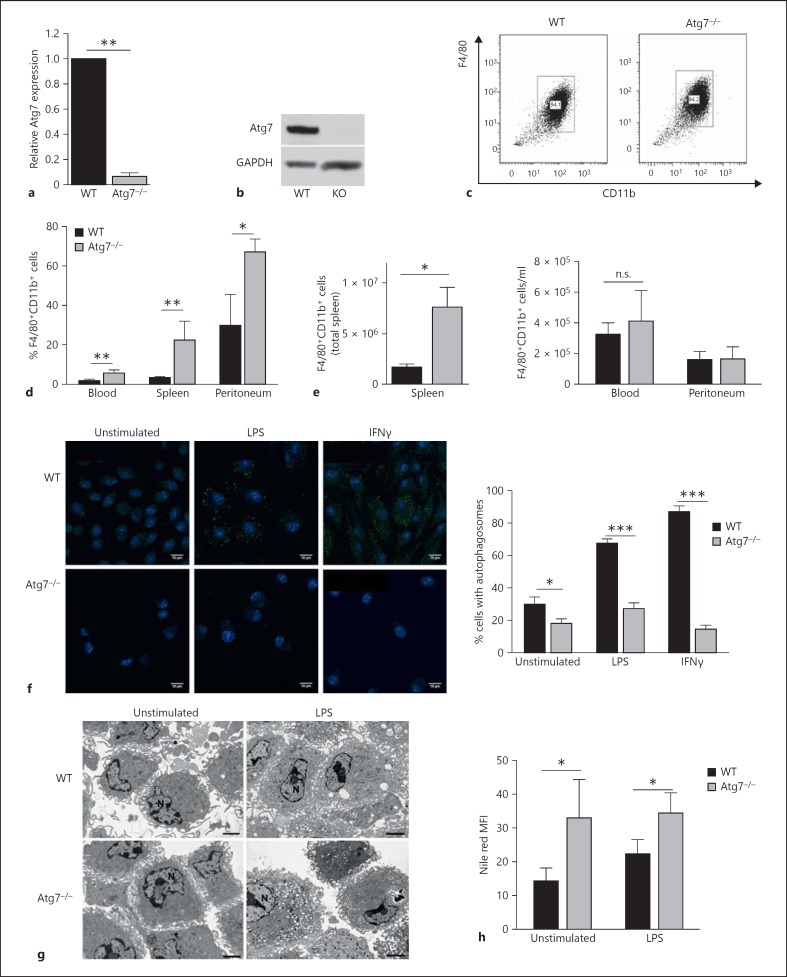
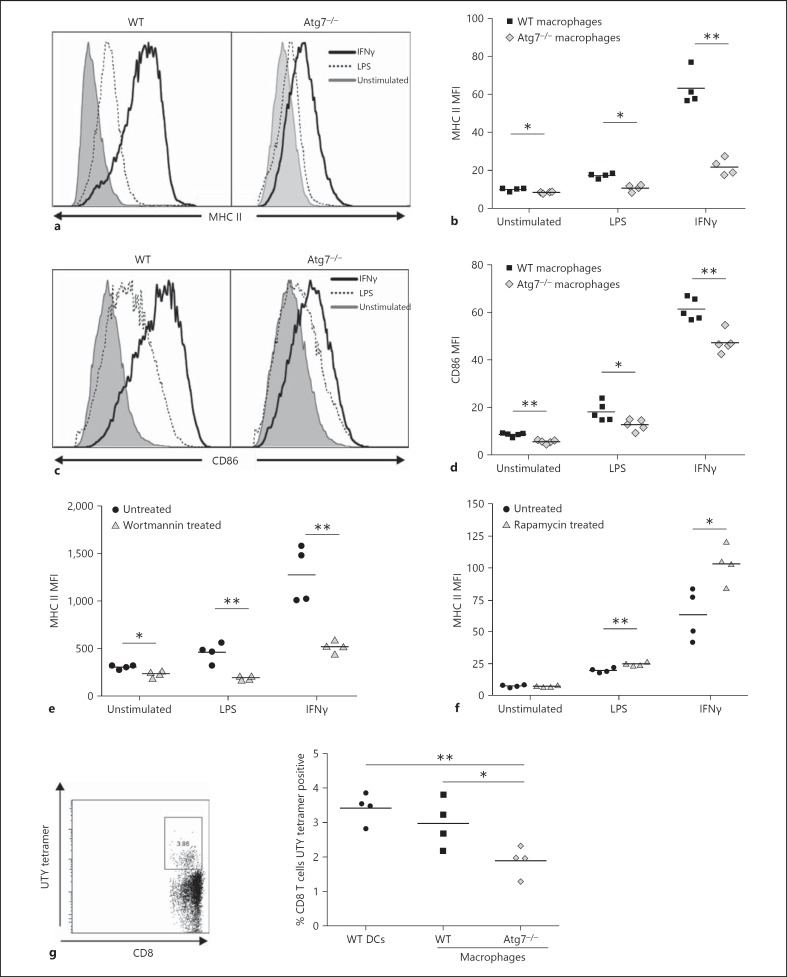
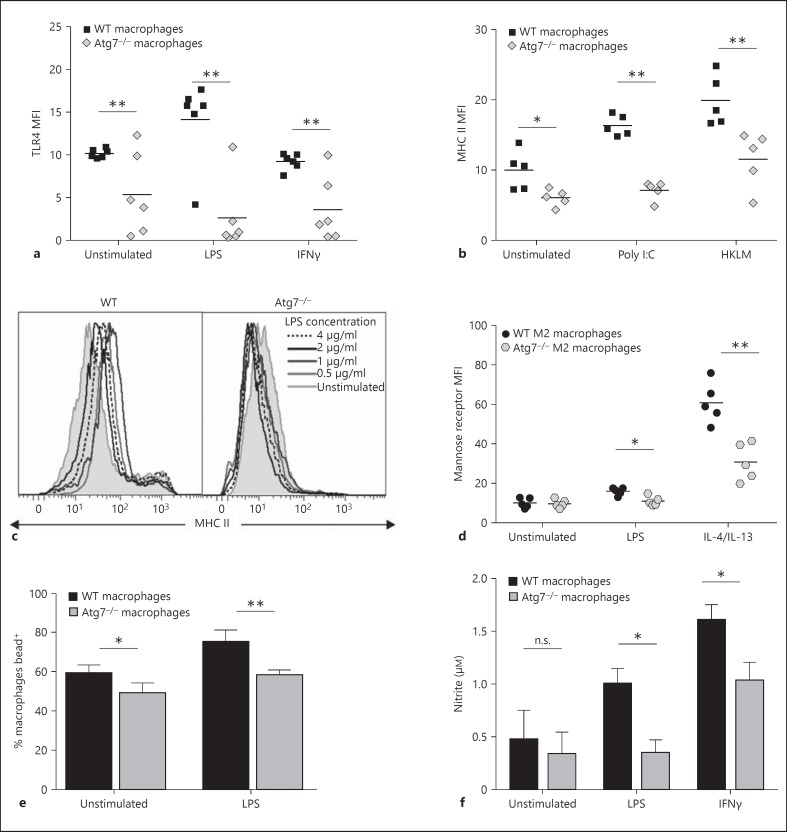
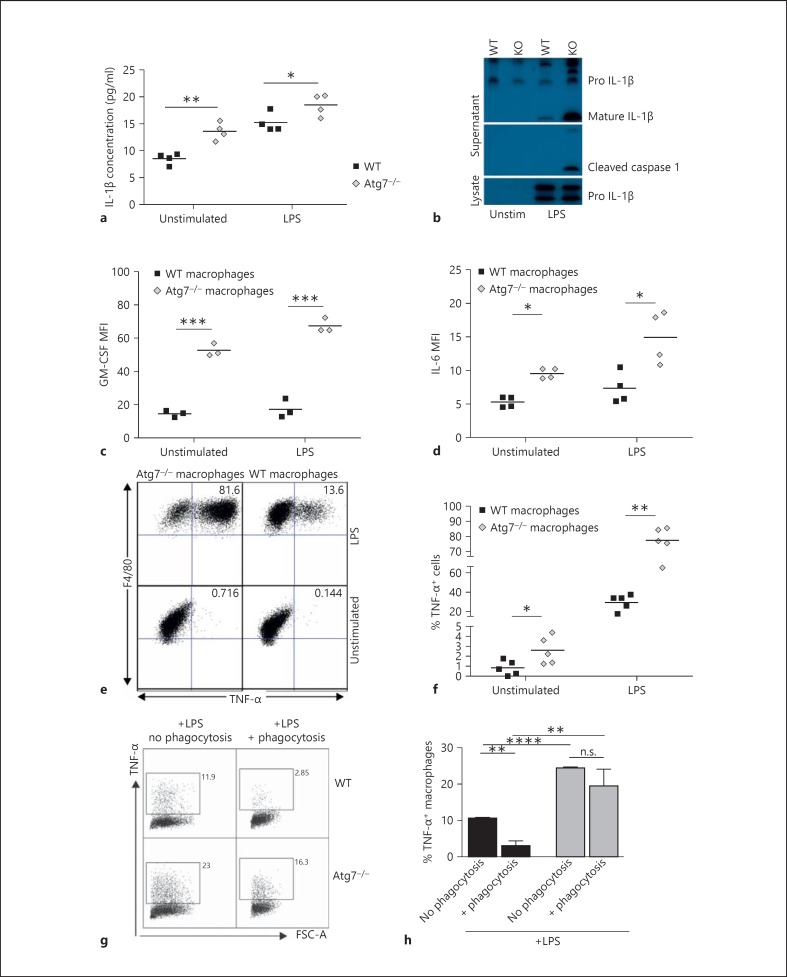
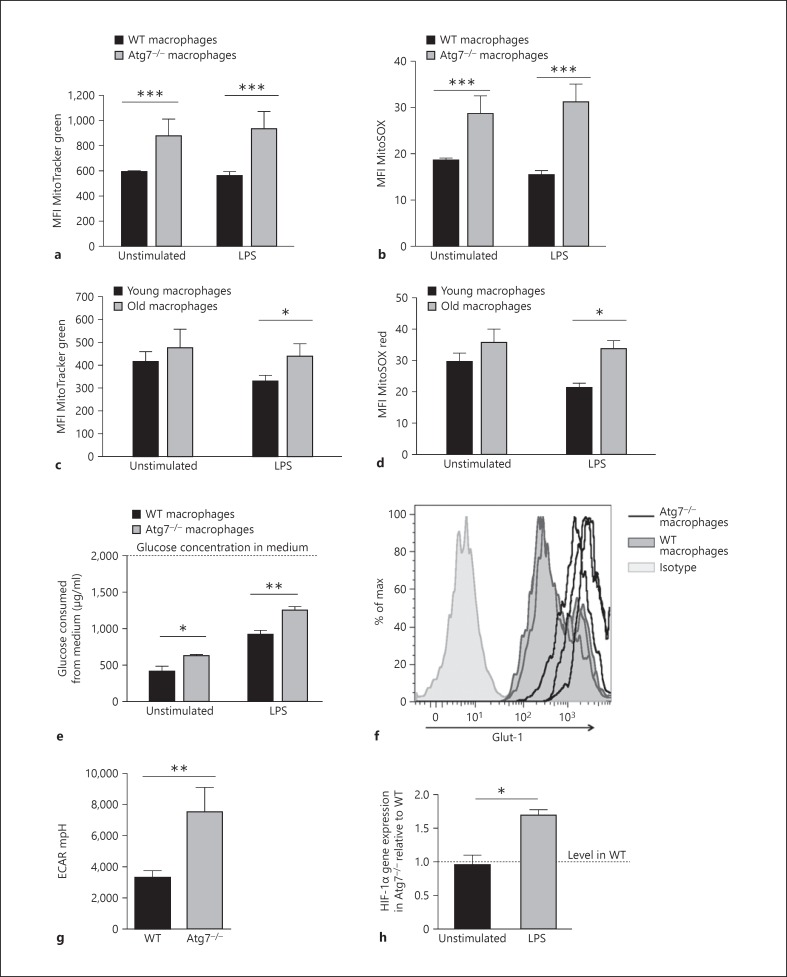
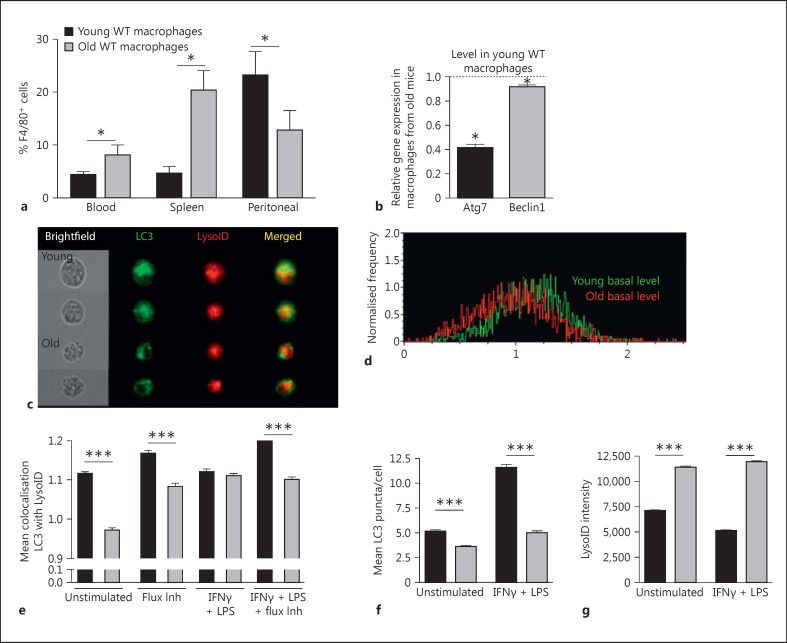
Macrophages provide a bridge linking innate and adaptive immunity. An increased frequency of macrophages and other myeloid cells paired with excessive cytokine production is commonly seen in the aging immune system, known as 'inflamm-aging'. It is presently unclear how healthy macrophages are maintained throughout life and what connects inflammation with myeloid dysfunction during aging. Autophagy, an intracellular degradation mechanism, has known links with aging and lifespan extension. Here, we show for the first time that autophagy regulates the acquisition of major aging features in macrophages. In the absence of the essential autophagy gene Atg7, macrophage populations are increased and key functions such as phagocytosis and nitrite burst are reduced, while the inflammatory cytokine response is significantly increased - a phenotype also observed in aged macrophages. Furthermore, reduced autophagy decreases surface antigen expression and skews macrophage metabolism toward glycolysis. We show that macrophages from aged mice exhibit significantly reduced autophagic flux compared to young mice. These data demonstrate that autophagy plays a critical role in the maintenance of macrophage homeostasis and function, regulating inflammation and metabolism and thereby preventing immunosenescence. Thus, autophagy modulation may prevent excess inflammation and preserve macrophage function during aging, improving immune responses and reducing the morbidity and mortality associated with inflamm-aging.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- T32 GM007309/GM/NIGMS NIH HHS/United States
- 17722/CRUK_/Cancer Research UK/United Kingdom
- MC_UU_12010/1/MRC_/Medical Research Council/United Kingdom
- BB/E021638/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/B/000C0409/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
