

A dynamin-actin interaction is required for vesicle scission during endocytosis in yeast
- PMID: 25772449
- PMCID: PMC4386032
- DOI: 10.1016/j.cub.2015.01.061
A dynamin-actin interaction is required for vesicle scission during endocytosis in yeast
Abstract
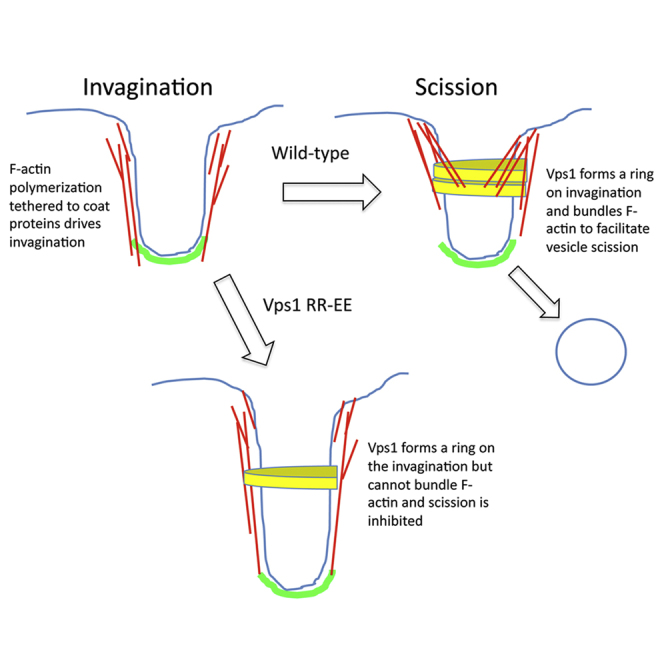
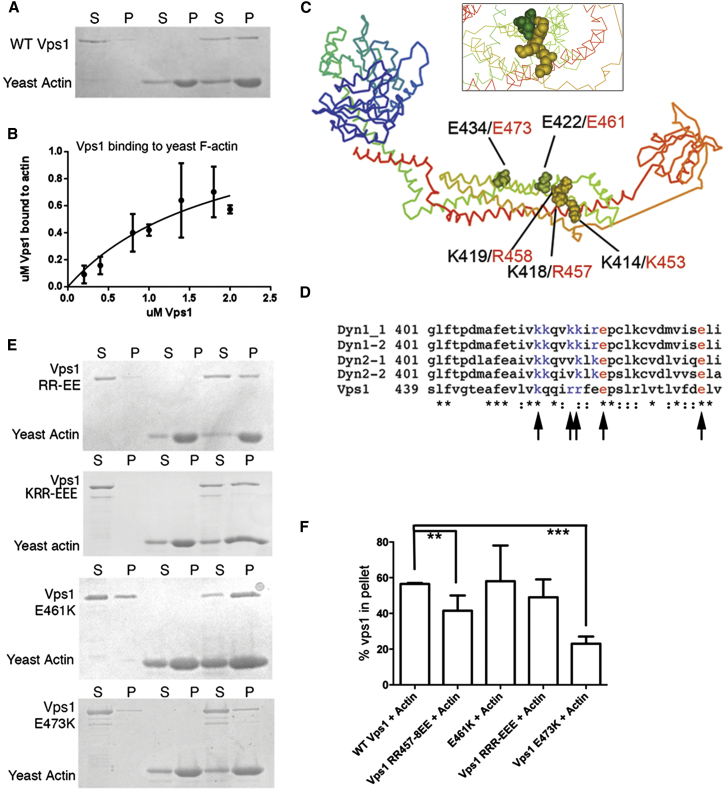
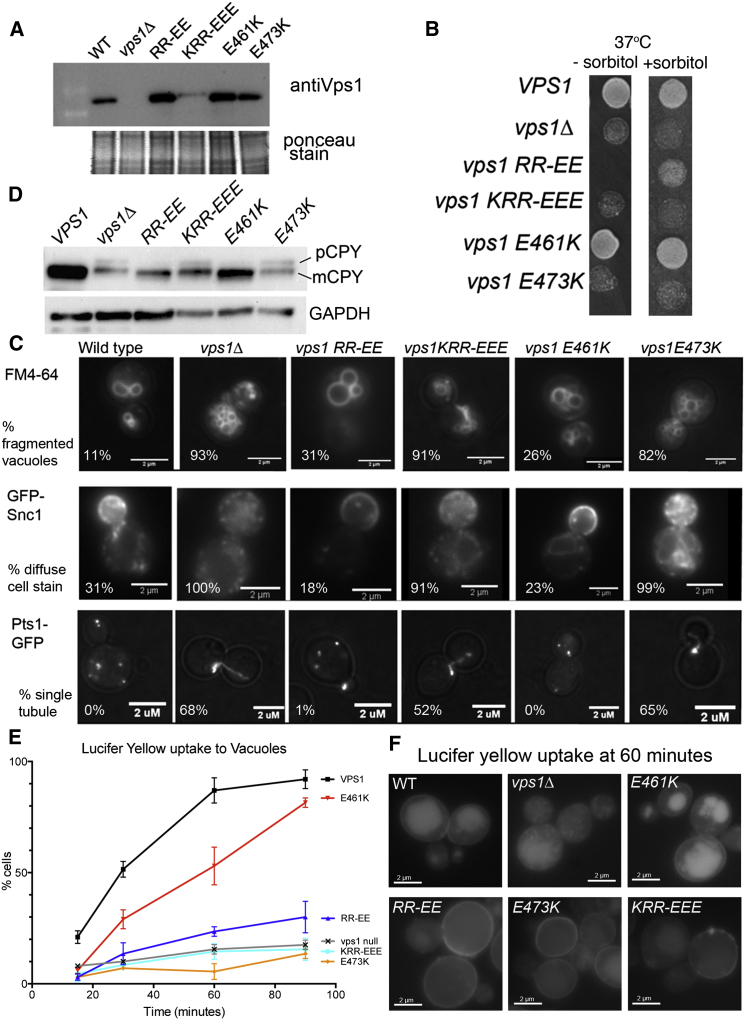
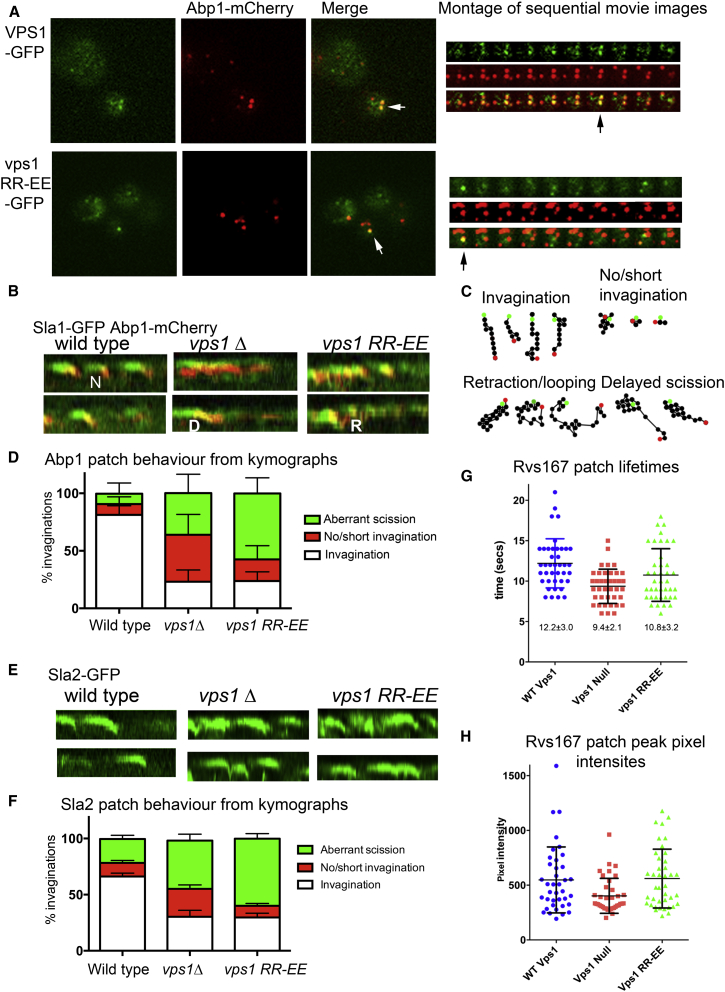
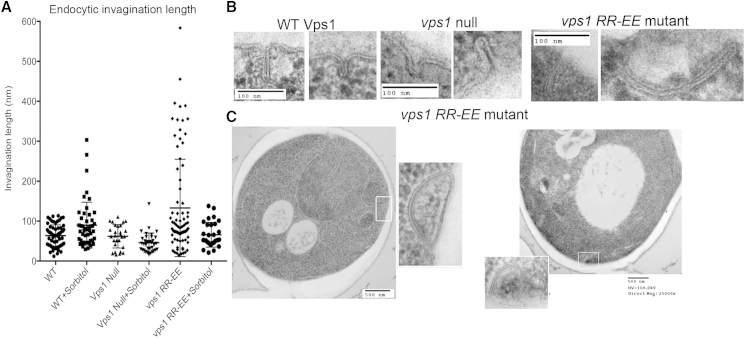
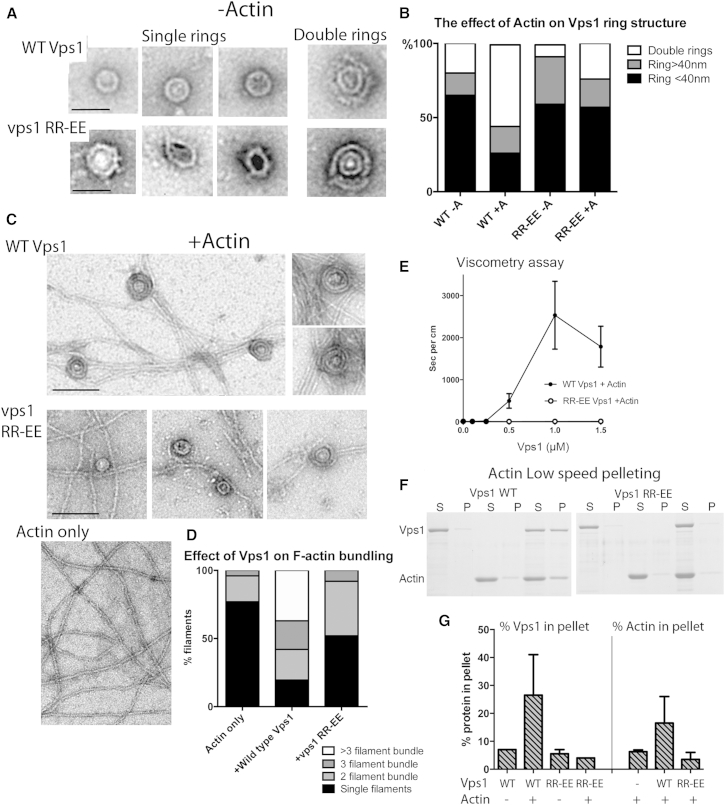
Actin is critical for endocytosis in yeast cells, and also in mammalian cells under tension. However, questions remain as to how force generated through actin polymerization is transmitted to the plasma membrane to drive invagination and scission. Here, we reveal that the yeast dynamin Vps1 binds and bundles filamentous actin. Mutational analysis of Vps1 in a helix of the stalk domain identifies a mutant RR457-458EE that binds actin more weakly. In vivo analysis of Vps1 function demonstrates that the mutation disrupts endocytosis but not other functions of Vps1 such as vacuolar trafficking or peroxisome fission. The mutant Vps1 is stably expressed in cells and co-localizes with the endocytic reporters Abp1 and the amphiphysin Rvs167. Detailed analysis of individual endocytic patch behavior indicates that the mutation causes aberrant movements in later stages of endocytosis, consistent with a scission defect. Ultrastructural analysis of yeast cells using electron microscopy reveals a significant increase in invagination depth, further supporting a role for the Vps1-actin interaction during scission. In vitro analysis of the mutant protein demonstrates that--like wild-type Vps1--it is able to form oligomeric rings, but, critically, it has lost its ability to bundle actin filaments into higher-order structures. A model is proposed in which actin filaments bind Vps1 during invagination, and this interaction is important to transduce the force of actin polymerization to the membrane to drive successful scission.
Copyright © 2015 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Urbanek A.N., Smith A.P., Allwood E.G., Booth W.I., Ayscough K.R. A novel actin-binding motif in Las17/WASP nucleates actin filaments independently of Arp2/3. Curr. Biol. 2013;23:196–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
