

Xenosurveillance: a novel mosquito-based approach for examining the human-pathogen landscape
- PMID: 25775236
- PMCID: PMC4361501
- DOI: 10.1371/journal.pntd.0003628
Xenosurveillance: a novel mosquito-based approach for examining the human-pathogen landscape
Abstract
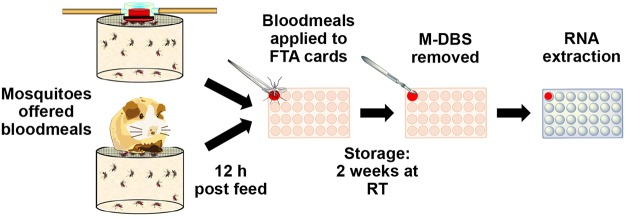
Background: Globally, regions at the highest risk for emerging infectious diseases are often the ones with the fewest resources. As a result, implementing sustainable infectious disease surveillance systems in these regions is challenging. The cost of these programs and difficulties associated with collecting, storing and transporting relevant samples have hindered them in the regions where they are most needed. Therefore, we tested the sensitivity and feasibility of a novel surveillance technique called xenosurveillance. This approach utilizes the host feeding preferences and behaviors of Anopheles gambiae, which are highly anthropophilic and rest indoors after feeding, to sample viruses in human beings. We hypothesized that mosquito bloodmeals could be used to detect vertebrate viral pathogens within realistic field collection timeframes and clinically relevant concentrations.
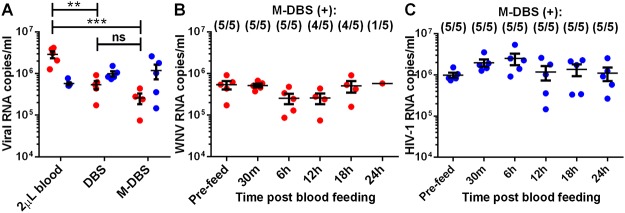
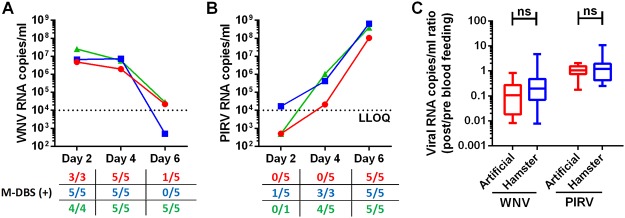
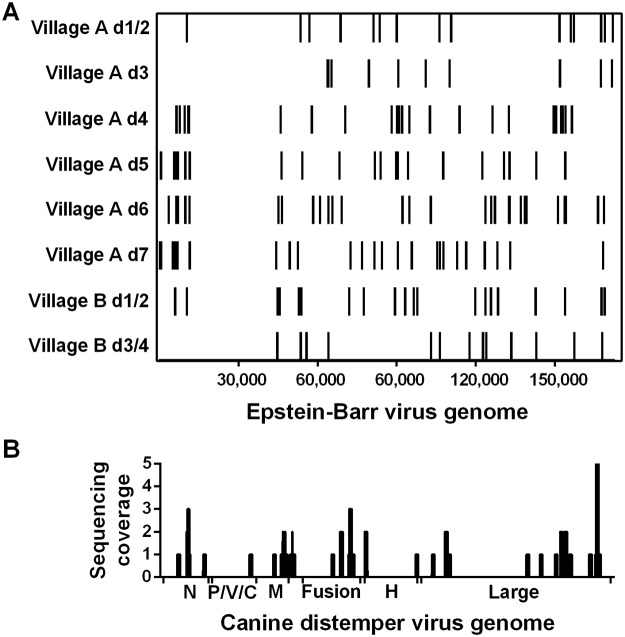
Methodology/principal findings: To validate this approach, we examined variables influencing virus detection such as the duration between mosquito blood feeding and mosquito processing, the pathogen nucleic acid stability in the mosquito gut and the pathogen load present in the host's blood at the time of bloodmeal ingestion using our laboratory model. Our findings revealed that viral nucleic acids, at clinically relevant concentrations, could be detected from engorged mosquitoes for up to 24 hours post feeding by qRT-PCR. Subsequently, we tested this approach in the field by examining blood from engorged mosquitoes from two field sites in Liberia. Using next-generation sequencing and PCR we were able to detect the genetic signatures of multiple viral pathogens including Epstein-Barr virus and canine distemper virus.
Conclusions/significance: Together, these data demonstrate the feasibility of xenosurveillance and in doing so validated a simple and non-invasive surveillance tool that could be used to complement current biosurveillance efforts.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Daszak P, Cunningham AA, Hyatt AD (2000) Emerging Infectious Diseases of Wildlife—Threats to Biodiversity and Human Health. Science 287: 443–449. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
