

Pathogenic mechanism of an autism-associated neuroligin mutation involves altered AMPA-receptor trafficking
- PMID: 25778475
- PMCID: PMC4573762
- DOI: 10.1038/mp.2015.20
Pathogenic mechanism of an autism-associated neuroligin mutation involves altered AMPA-receptor trafficking
Abstract
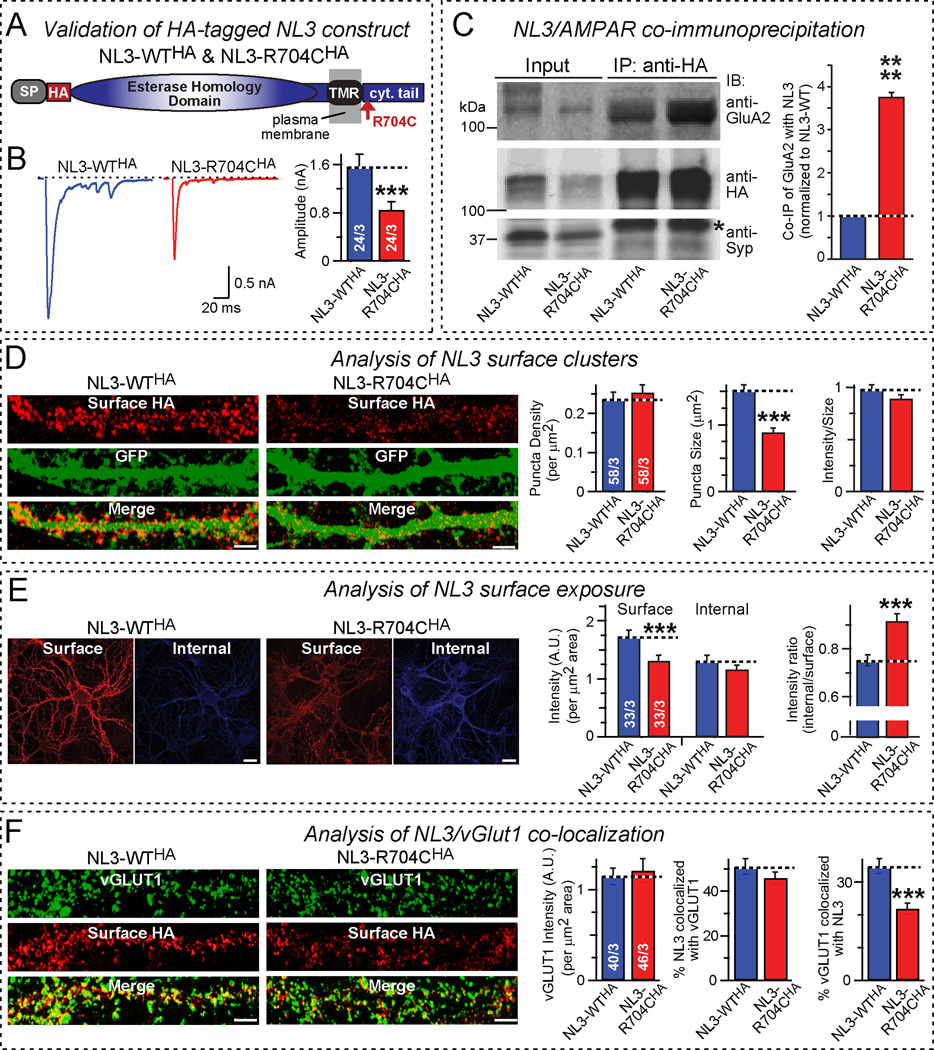
Neuroligins are postsynaptic cell-adhesion molecules that bind to presynaptic neurexins. Although the general synaptic role of neuroligins is undisputed, their specific functions at a synapse remain unclear, even controversial. Moreover, many neuroligin gene mutations were associated with autism, but the pathophysiological relevance of these mutations is often unknown, and their mechanisms of action uninvestigated. Here, we examine the synaptic effects of an autism-associated neuroligin-4 substitution (called R704C), which mutates a cytoplasmic arginine residue that is conserved in all neuroligins. We show that the R704C mutation, when introduced into neuroligin-3, enhances the interaction between neuroligin-3 and AMPA receptors, increases AMPA-receptor internalization and decreases postsynaptic AMPA-receptor levels. When introduced into neuroligin-4, conversely, the R704C mutation unexpectedly elevated AMPA-receptor-mediated synaptic responses. These results suggest a general functional link between neuroligins and AMPA receptors, indicate that both neuroligin-3 and -4 act at excitatory synapses but perform surprisingly distinct functions, and demonstrate that the R704C mutation significantly impairs the normal function of neuroligin-4, thereby validating its pathogenicity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
