

Differential regulation of proton-sensitive ion channels by phospholipids: a comparative study between ASICs and TRPV1
- PMID: 25781982
- PMCID: PMC4362947
- DOI: 10.1371/journal.pone.0122014
Differential regulation of proton-sensitive ion channels by phospholipids: a comparative study between ASICs and TRPV1
Abstract
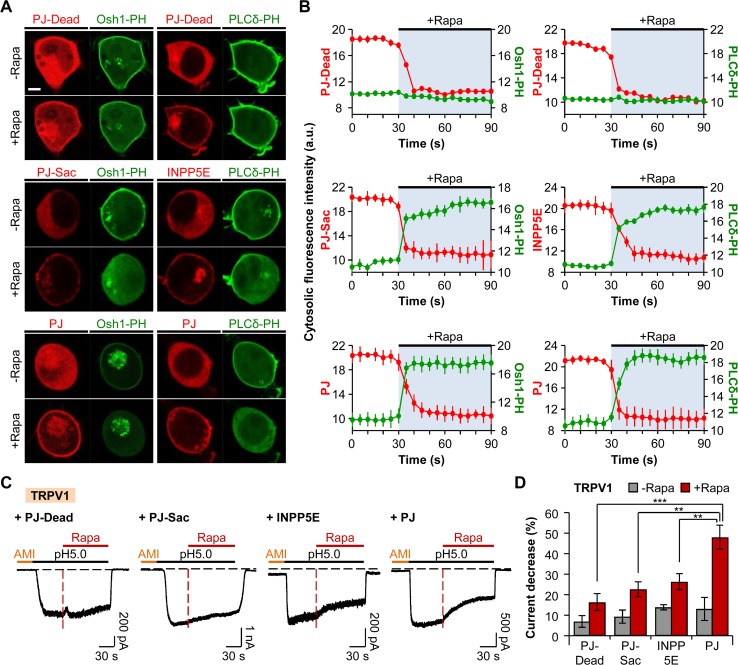
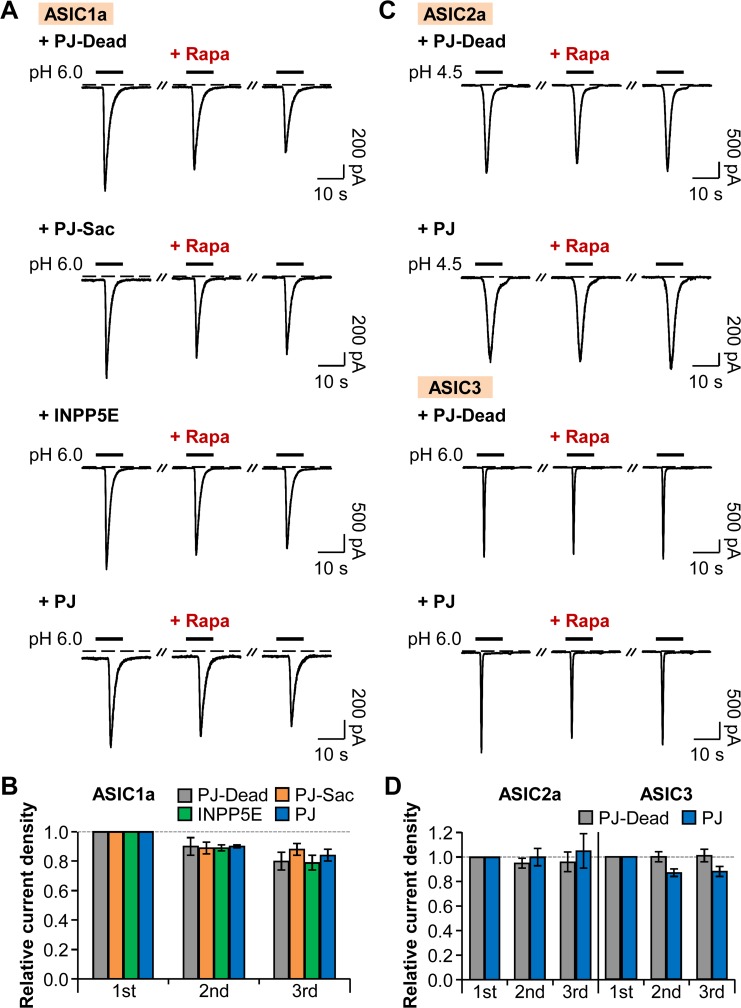
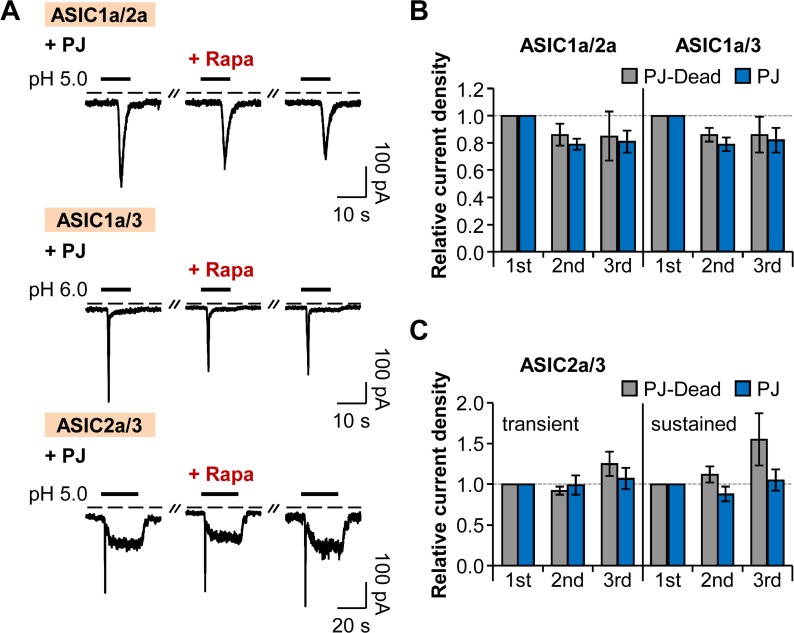
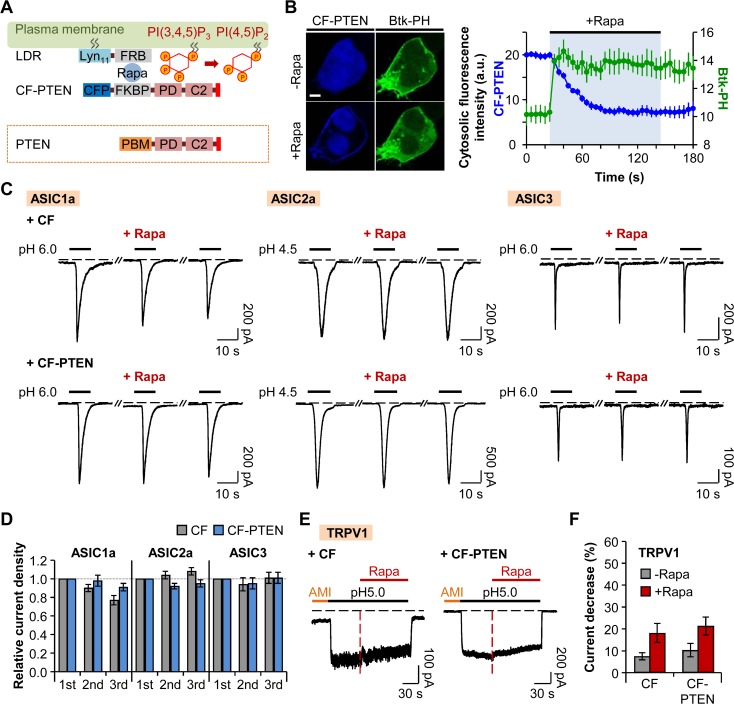
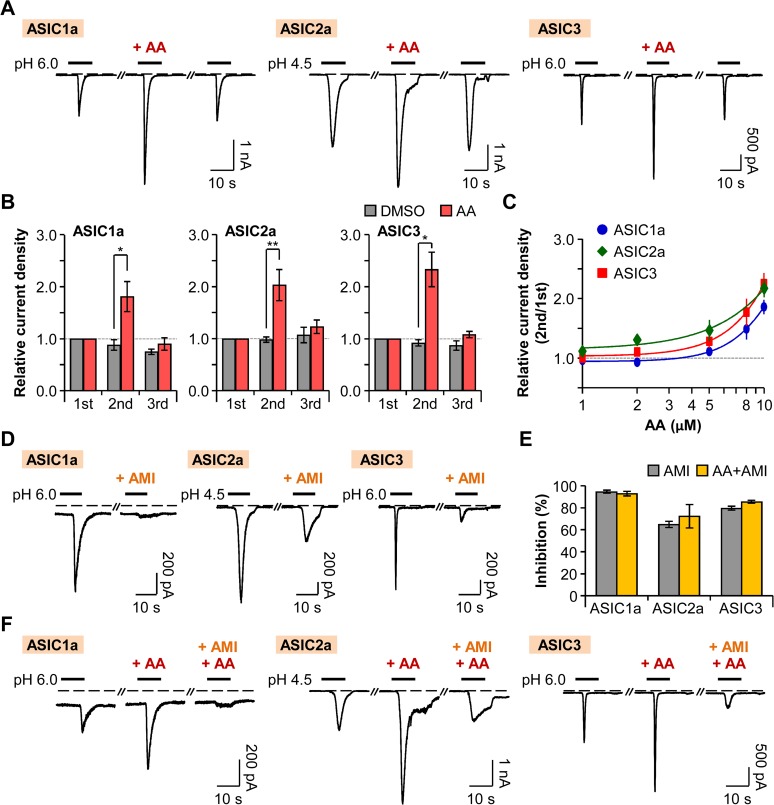
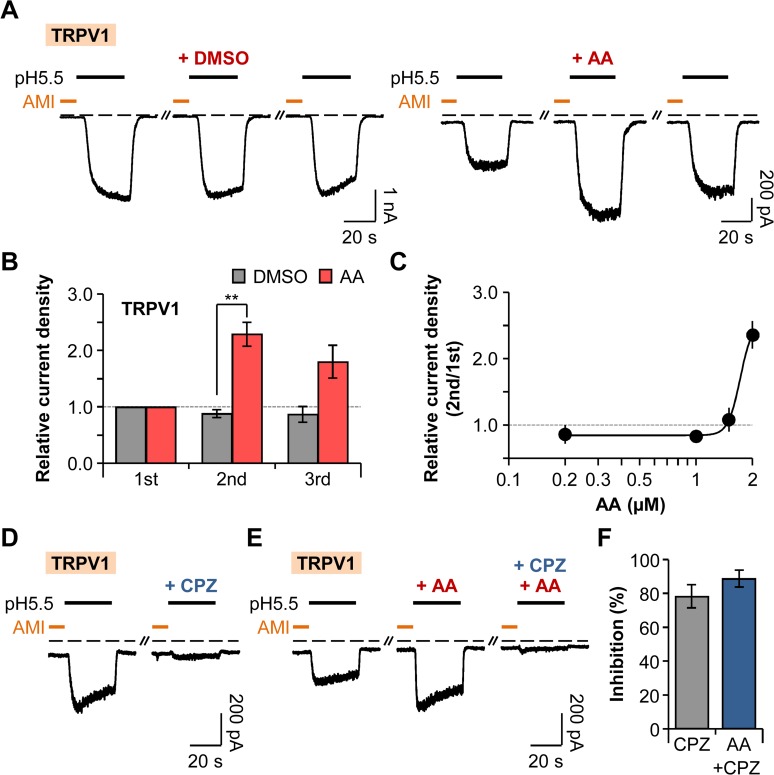
Protons are released in pain-generating pathological conditions such as inflammation, ischemic stroke, infection, and cancer. During normal synaptic activities, protons are thought to play a role in neurotransmission processes. Acid-sensing ion channels (ASICs) are typical proton sensors in the central nervous system (CNS) and the peripheral nervous system (PNS). In addition to ASICs, capsaicin- and heat-activated transient receptor potential vanilloid 1 (TRPV1) channels can also mediate proton-mediated pain signaling. In spite of their importance in perception of pH fluctuations, the regulatory mechanisms of these proton-sensitive ion channels still need to be further investigated. Here, we compared regulation of ASICs and TRPV1 by membrane phosphoinositides, which are general cofactors of many receptors and ion channels. We observed that ASICs do not require membrane phosphatidylinositol 4-phosphate (PI(4)P) or phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) for their function. However, TRPV1 currents were inhibited by simultaneous breakdown of PI(4)P and PI(4,5)P2. By using a novel chimeric protein, CF-PTEN, that can specifically dephosphorylate at the D3 position of phosphatidylinositol 3,4,5-trisphosphate (PI(3,4,5)P3), we also observed that neither ASICs nor TRPV1 activities were altered by depletion of PI(3,4,5)P3 in intact cells. Finally, we compared the effects of arachidonic acid (AA) on two proton-sensitive ion channels. We observed that AA potentiates the currents of both ASICs and TRPV1, but that they have different recovery aspects. In conclusion, ASICs and TRPV1 have different sensitivities toward membrane phospholipids, such as PI(4)P, PI(4,5)P2, and AA, although they have common roles as proton sensors. Further investigation about the complementary roles and respective contributions of ASICs and TRPV1 in proton-mediated signaling is necessary.
Conflict of interest statement
Figures
Similar articles
-
Morphine inhibits acid-sensing ion channel currents in rat dorsal root ganglion neurons.Brain Res. 2014 Mar 20;1554:12-20. doi: 10.1016/j.brainres.2014.01.042. Epub 2014 Feb 1. Brain Res. 2014. PMID: 24491633
-
Activation of TRPV1 channels inhibits mechanosensitive Piezo channel activity by depleting membrane phosphoinositides.Sci Signal. 2015 Feb 10;8(363):ra15. doi: 10.1126/scisignal.2005667. Sci Signal. 2015. PMID: 25670203 Free PMC article.
-
PAR-2 activation enhances weak acid-induced ATP release through TRPV1 and ASIC sensitization in human esophageal epithelial cells.Am J Physiol Gastrointest Liver Physiol. 2015 Oct 15;309(8):G695-702. doi: 10.1152/ajpgi.00162.2015. Epub 2015 Aug 20. Am J Physiol Gastrointest Liver Physiol. 2015. PMID: 26294672
-
Acid-sensing ion channels: dual function proteins for chemo-sensing and mechano-sensing.J Biomed Sci. 2018 May 24;25(1):46. doi: 10.1186/s12929-018-0448-y. J Biomed Sci. 2018. PMID: 29793480 Free PMC article. Review.
-
The function and regulation of acid-sensing ion channels (ASICs) and the epithelial Na(+) channel (ENaC): IUPHAR Review 19.Br J Pharmacol. 2016 Sep;173(18):2671-701. doi: 10.1111/bph.13533. Epub 2016 Aug 10. Br J Pharmacol. 2016. PMID: 27278329 Free PMC article. Review.
Cited by
-
Acid-sensing ion channel 1a activates IKCa/SKCa channels and contributes to endothelium-dependent dilation.J Gen Physiol. 2023 Feb 6;155(2):e202213173. doi: 10.1085/jgp.202213173. Epub 2022 Dec 9. J Gen Physiol. 2023. PMID: 36484717 Free PMC article.
-
Harnessing the Therapeutic Potential of Capsaicin and Its Analogues in Pain and Other Diseases.Molecules. 2016 Jul 23;21(8):966. doi: 10.3390/molecules21080966. Molecules. 2016. PMID: 27455231 Free PMC article. Review.
-
ASIC2a-dependent increase of ASIC3 surface expression enhances the sustained component of the currents.BMB Rep. 2016 Oct;49(10):542-547. doi: 10.5483/bmbrep.2016.49.10.057. BMB Rep. 2016. PMID: 27241858 Free PMC article.
-
Rapid resensitization of ASIC2a is conferred by three amino acid residues in the N terminus.J Gen Physiol. 2019 Jul 1;151(7):944-953. doi: 10.1085/jgp.201812224. Epub 2019 Apr 22. J Gen Physiol. 2019. PMID: 31010811 Free PMC article.
-
Endothelin-1 enhances acid-sensing ion channel currents in rat primary sensory neurons.Acta Pharmacol Sin. 2020 Aug;41(8):1049-1057. doi: 10.1038/s41401-019-0348-z. Epub 2020 Feb 27. Acta Pharmacol Sin. 2020. PMID: 32107467 Free PMC article.
References
-
- Smith ML, von Hanwehr R, Siesjö BK. Changes in extra- and intracellular pH in the brain during and following ischemia in hyperglycemic and in moderately hypoglycemic rats. J Cereb Blood Flow Metab. 1986;6(5): 574–83. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
