

A Residue Quartet in the Extracellular Domain of the Prolactin Receptor Selectively Controls Mitogen-activated Protein Kinase Signaling
- PMID: 25784554
- PMCID: PMC4424329
- DOI: 10.1074/jbc.M115.639096
A Residue Quartet in the Extracellular Domain of the Prolactin Receptor Selectively Controls Mitogen-activated Protein Kinase Signaling
Abstract
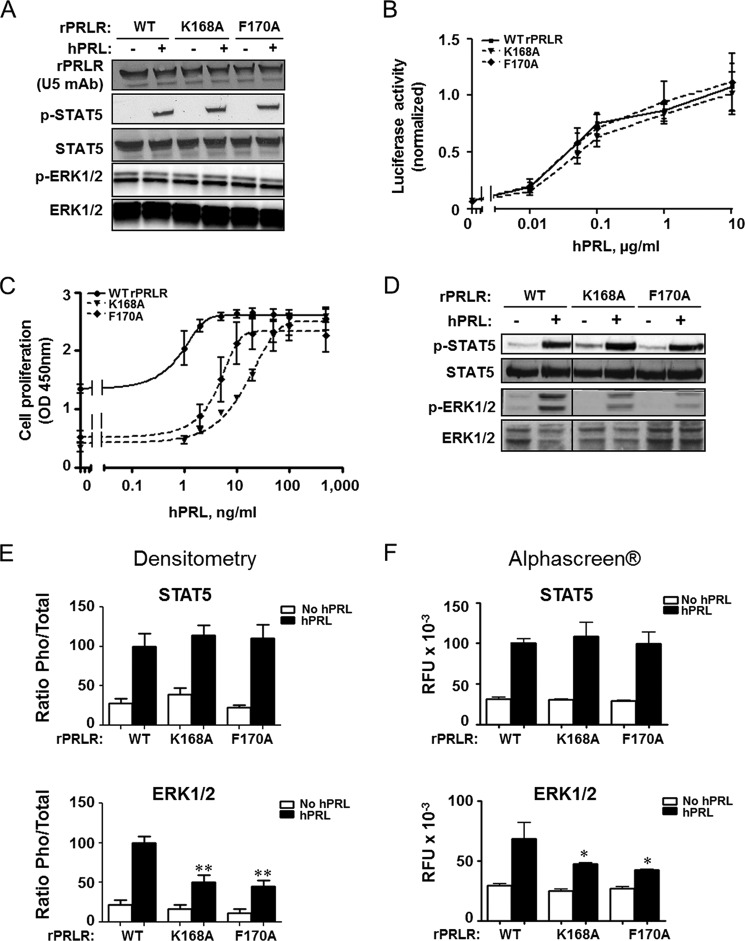
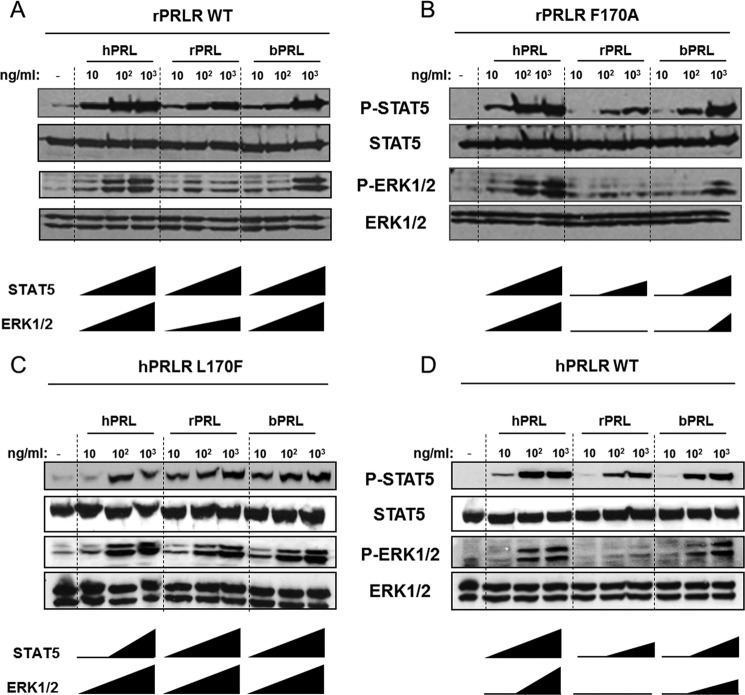
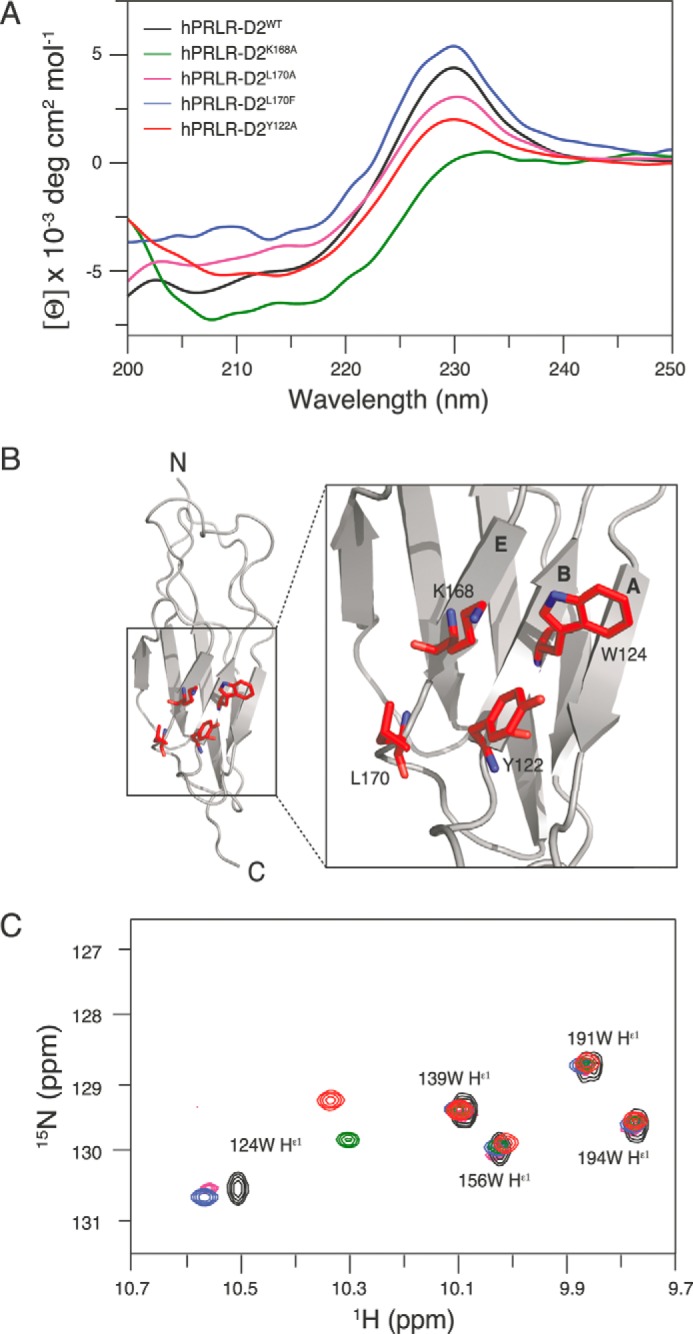
Cytokine receptors elicit several signaling pathways, but it is poorly understood how they select and discriminate between them. We have scrutinized the prolactin receptor as an archetype model of homodimeric cytokine receptors to address the role of the extracellular membrane proximal domain in signal transfer and pathway selection. Structure-guided manipulation of residues involved in the receptor dimerization interface identified one residue (position 170) that in cell-based assays profoundly altered pathway selectivity and species-specific bio-characteristics. Subsequent in vitro spectroscopic and nuclear magnetic resonance analyses revealed that this residue was part of a residue quartet responsible for specific local structural changes underlying these effects. This included alteration of a novel aromatic T-stack within the membrane proximal domain, which promoted selective signaling affecting primarily the MAPK (ERK1/2) pathway. Importantly, activation of the MAPK pathway correlated with in vitro stabilities of ternary ligand·receptor complexes, suggesting a threshold mean lifetime of the complex necessary to achieve maximal activation. No such dependence was observed for STAT5 signaling. Thus, this study establishes a residue quartet in the extracellular membrane proximal domain of homodimeric cytokine receptors as a key regulator of intracellular signaling discrimination.
Keywords: Cell Proliferation; Cell Surface Receptor; Cytokine; Extracellular Signal-regulated Kinase (ERK); Nuclear Magnetic Resonance (NMR); STAT Transcription Factor.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures







References
-
- Karnitz L. M., Abraham R. T. (1995) Cytokine receptor signaling mechanisms. Curr. Opin. Immunol. 7, 320–326 - PubMed
-
- Chang F., Steelman L. S., Lee J. T., Shelton J. G., Navolanic P. M., Blalock W. L., Franklin R. A., McCubrey J. A. (2003) Signal transduction mediated by the Ras/Raf/MEK/ERK pathway from cytokine receptors to transcription factors: potential targeting for therapeutic intervention. Leukemia 17, 1263–1293 - PubMed
-
- Li W., Lan H., Liu H., Fu Z., Yang Y., Han W., Guo F., Liu Y., Zhang H., Liu J., Zheng X. (2013) The activation and differential signalling of the growth hormone receptor induced by pGH or anti-idiotypic monoclonal antibodies in primary rat hepatocytes. Mol. Cell. Endocrinol. 376, 51–59 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
