

The c-Cbl ubiquitin ligase regulates nuclear β-catenin and angiogenesis by its tyrosine phosphorylation mediated through the Wnt signaling pathway
- PMID: 25784557
- PMCID: PMC4432275
- DOI: 10.1074/jbc.M114.616623
The c-Cbl ubiquitin ligase regulates nuclear β-catenin and angiogenesis by its tyrosine phosphorylation mediated through the Wnt signaling pathway
Abstract
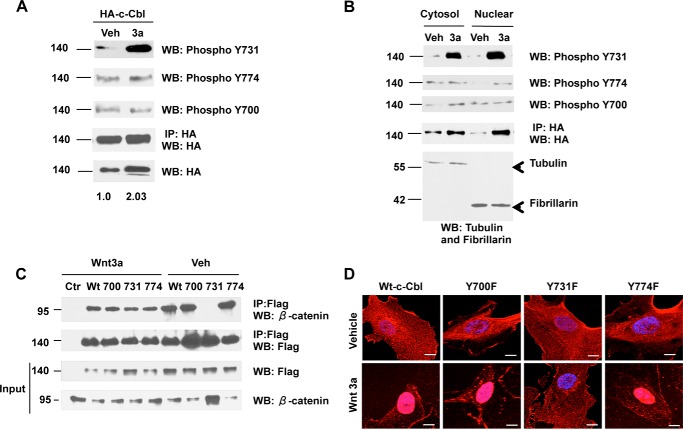
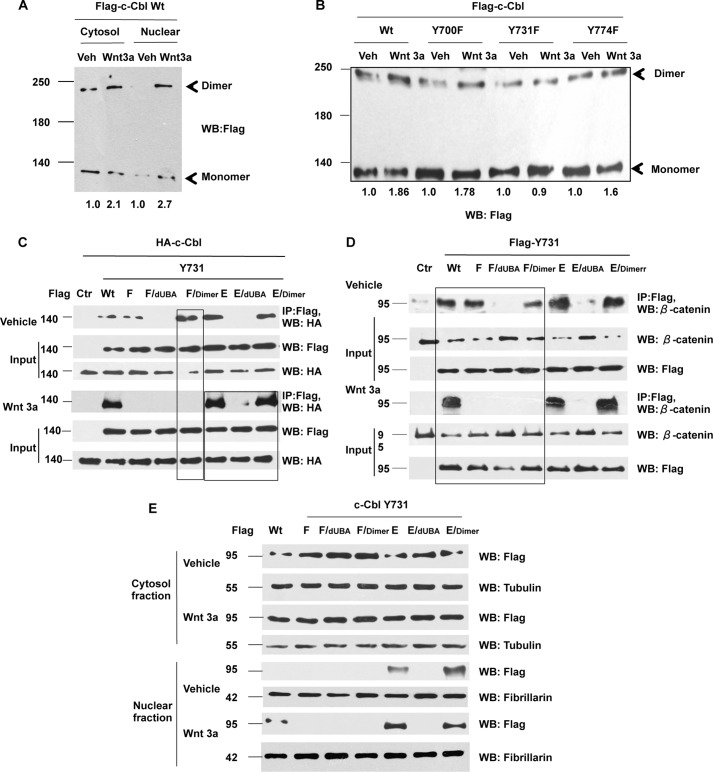
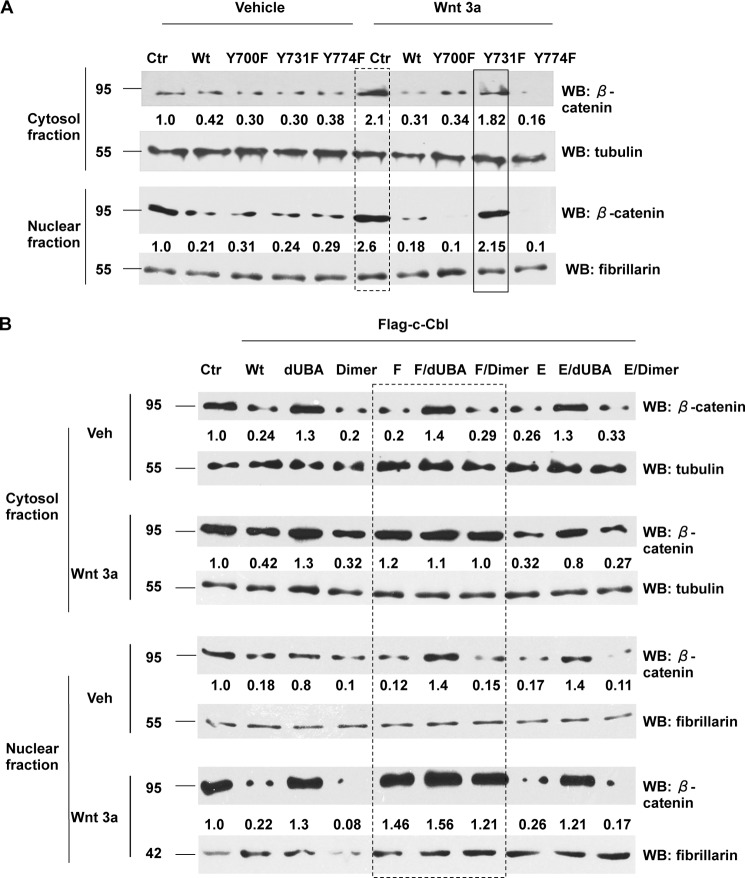
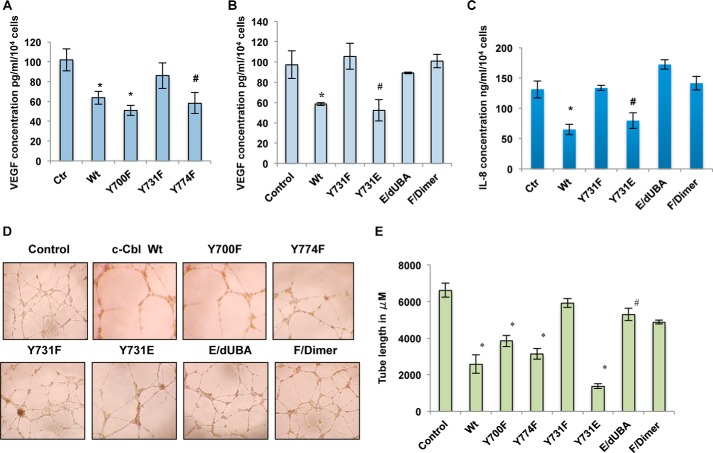
Wnt signaling plays important roles in both the tumor-induced angiogenesis and tumorigenesis through the transcriptionally active nuclear β-catenin. Recently, c-Cbl was identified as a unique E3 ubiquitin ligase targeting the active nuclear β-catenin. However, little is known about the molecular mechanisms by which c-Cbl regulates ubiquitination and degradation of active β-catenin. Here, we demonstrate that Wnt activation promotes the phosphorylation of c-Cbl at tyrosine 731(Tyr-731), which increases c-Cbl dimerization and binding to β-catenin. Tyr-731 phosphorylation and dimerization mediate c-Cbl nuclear translocation and lead to the degradation of nuclearly active β-catenin in the Wnt-on phase. c-Cbl activation also inhibits expression of the pro-angiogenic Wnt targets, IL-8 and VEGF. Phospho-Tyr-731-inactive mutant c-Cbl (Y731F) enhances and phosphomimetic mutant c-Cbl (Y731E) suppresses angiogenesis in zebrafish. Taken together, we have identified a novel mechanism for the regulation of active nuclear β-catenin by c-Cbl and its critical role in angiogenesis. This mechanism can be further explored to modulate both the pathological angiogenesis and the tumorigenesis.
Keywords: Angiogenesis; Cbl; E3 Ubiquitin Ligase; Tumor Cell Biology; Tumorigenesis; Tumors; Wnt Signaling; Zebrafish; β-Catenin.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
c-Cbl, a ubiquitin E3 ligase that targets active β-catenin: a novel layer of Wnt signaling regulation.J Biol Chem. 2013 Aug 9;288(32):23505-17. doi: 10.1074/jbc.M113.473801. Epub 2013 Jun 6. J Biol Chem. 2013. PMID: 23744067 Free PMC article.
-
c-Cbl Expression Correlates with Human Colorectal Cancer Survival and Its Wnt/β-Catenin Suppressor Function Is Regulated by Tyr371 Phosphorylation.Am J Pathol. 2018 Aug;188(8):1921-1933. doi: 10.1016/j.ajpath.2018.05.007. Epub 2018 Jul 17. Am J Pathol. 2018. PMID: 30029779 Free PMC article. Clinical Trial.
-
c-Cbl: An Important Regulator and a Target in Angiogenesis and Tumorigenesis.Cells. 2019 May 23;8(5):498. doi: 10.3390/cells8050498. Cells. 2019. PMID: 31126146 Free PMC article. Review.
-
The ciliary protein nephrocystin-4 translocates the canonical Wnt regulator Jade-1 to the nucleus to negatively regulate β-catenin signaling.J Biol Chem. 2012 Jul 20;287(30):25370-80. doi: 10.1074/jbc.M112.385658. Epub 2012 May 31. J Biol Chem. 2012. PMID: 22654112 Free PMC article.
-
E3 ubiquitin ligase-mediated regulation of bone formation and tumorigenesis.Cell Death Dis. 2013 Jan 17;4(1):e463. doi: 10.1038/cddis.2012.217. Cell Death Dis. 2013. PMID: 23328670 Free PMC article. Review.
Cited by
-
Transcriptional Regulation of Wnt/β-Catenin Pathway in Colorectal Cancer.Cells. 2020 Sep 19;9(9):2125. doi: 10.3390/cells9092125. Cells. 2020. PMID: 32961708 Free PMC article. Review.
-
Ubiquitin Ligases Involved in the Regulation of Wnt, TGF-β, and Notch Signaling Pathways and Their Roles in Mouse Development and Homeostasis.Genes (Basel). 2019 Oct 16;10(10):815. doi: 10.3390/genes10100815. Genes (Basel). 2019. PMID: 31623112 Free PMC article. Review.
-
Multifunctional DDX3: dual roles in various cancer development and its related signaling pathways.Am J Cancer Res. 2016 Jan 15;6(2):387-402. eCollection 2016. Am J Cancer Res. 2016. PMID: 27186411 Free PMC article. Review.
-
CTEN Inhibits Tumor Angiogenesis and Growth by Targeting VEGFA Through Down-Regulation of β-Catenin in Breast Cancer.Technol Cancer Res Treat. 2021 Jan-Dec;20:15330338211045506. doi: 10.1177/15330338211045506. Technol Cancer Res Treat. 2021. PMID: 34817293 Free PMC article.
-
Breakdown of Filamentous Myofibrils by the UPS-Step by Step.Biomolecules. 2021 Jan 15;11(1):110. doi: 10.3390/biom11010110. Biomolecules. 2021. PMID: 33467597 Free PMC article. Review.
References
-
- Weis S. M., Cheresh D. A. (2011) Tumor angiogenesis: molecular pathways and therapeutic targets. Nat. Med. 17, 1359–1370 - PubMed
-
- Reis M., Liebner S. (2013) Wnt signaling in the vasculature. Exp. Cell Res. 319, 1317–1323 - PubMed
-
- Nusse R. (2005) Wnt signaling in disease and in development. Cell Res. 15, 28–32 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
