

Consequence of the tumor-associated conversion to cyclin D1b
- PMID: 25787974
- PMCID: PMC4492821
- DOI: 10.15252/emmm.201404242
Consequence of the tumor-associated conversion to cyclin D1b
Abstract
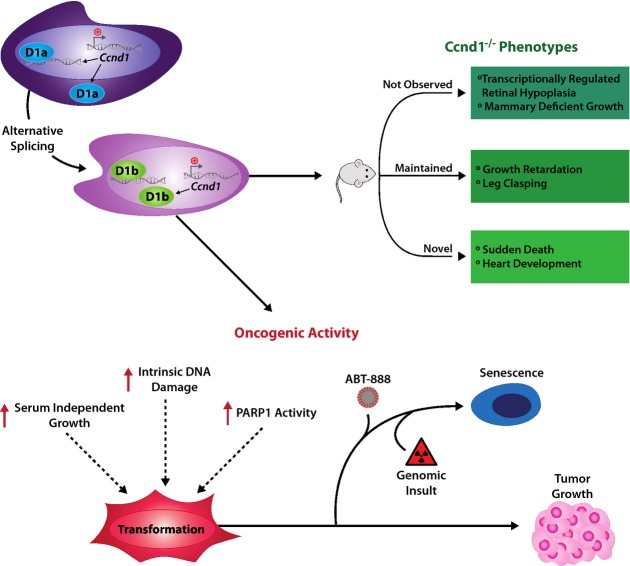
Clinical evidence suggests that cyclin D1b, a variant of cyclin D1, is associated with tumor progression and poor outcome. However, the underlying molecular basis was unknown. Here, novel models were created to generate a genetic switch from cyclin D1 to cyclin D1b. Extensive analyses uncovered overlapping but non-redundant functions of cyclin D1b compared to cyclin D1 on developmental phenotypes, and illustrated the importance of the transcriptional regulatory functions of cyclin D1b in vivo. Data obtained identify cyclin D1b as an oncogene, wherein cyclin D1b expression under the endogenous promoter induced cellular transformation and further cooperated with known oncogenes to promote tumor growth in vivo. Further molecular interrogation uncovered unexpected links between cyclin D1b and the DNA damage/PARP1 regulatory networks, which could be exploited to suppress cyclin D1b-driven tumors. Collectively, these data are the first to define the consequence of cyclin D1b expression on normal cellular function, present evidence for cyclin D1b as an oncogene, and provide pre-clinical evidence of effective methods to thwart growth of cells dependent upon this oncogenic variant.
Keywords: PARP; cell cycle; cyclin; cyclin D1b.
© 2015 The Authors. Published under the terms of the CC BY 4.0 license.
Figures
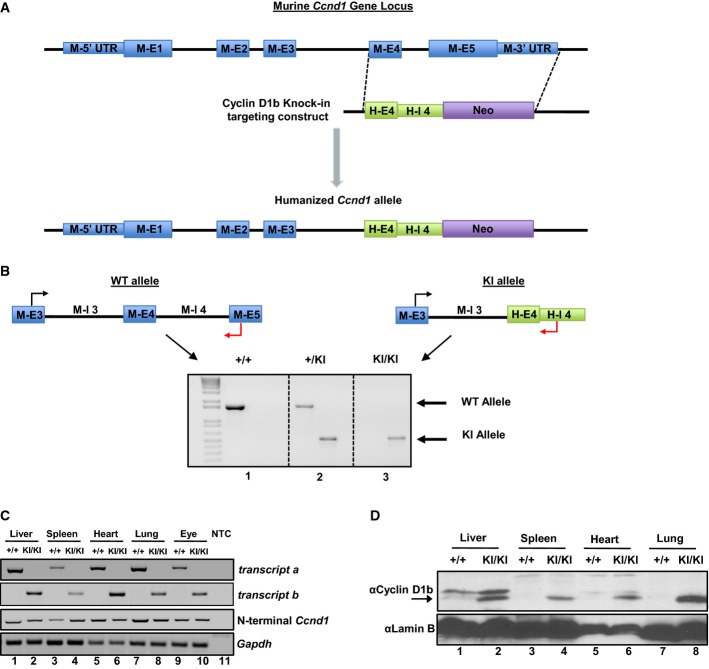
Representative schematic of the targeting construct generated to humanize the Ccnd1 exon 4/5 genomic locus to produce cyclin D1b.
Top: Schematic of primer pairs designed to discriminate between wild-type and knock-in alleles. Bottom: Representative genotyping of Ccnd1+/+, Ccnd1+/KI, and Ccnd1KI/KI mice validating specificity of the primer pairs and somatic insertion of the targeting construct.
PCR analysis of transcript b expression in organs harvested from Ccnd1+/+ and Ccnd1KI/KI mice, demonstrating production of transcript b specifically in KI mice. Gapdh serves as a control (NTC, Non-template control).
Immunoblot from parallel samples in (C) utilizing antisera specific to the 33 amino acids generated by human CCND1 intron 4. Lamin B serves as a control. Arrow indicates the cyclin D1b band.
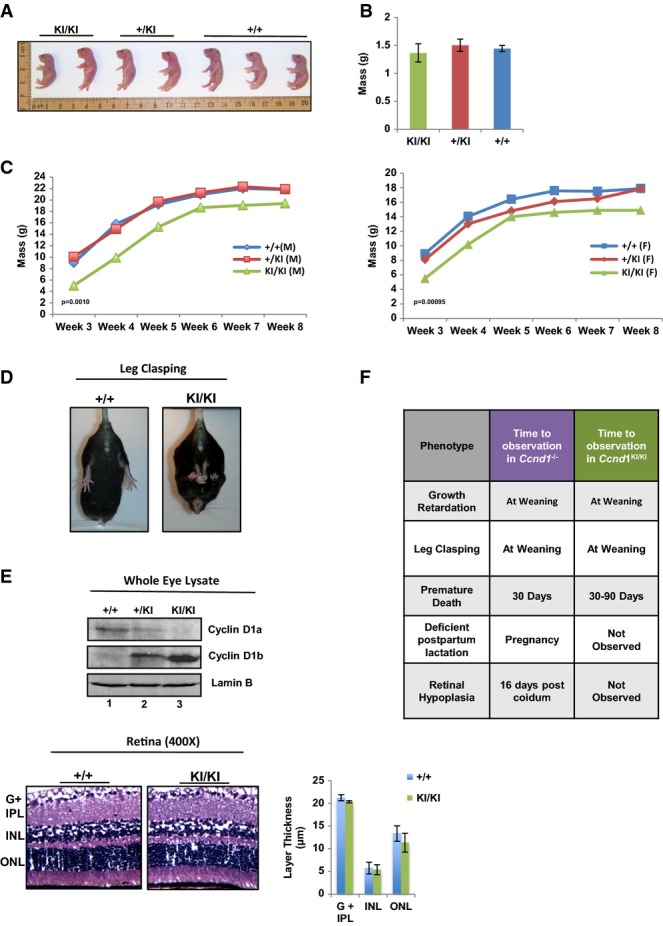
Neonates from 2 independent litters of Ccnd1+/KI × Ccnd1+/KI crosses were sacrificed at birth and genotyped. Mice were organized by genotype and total size measured.
Mass of neonates was calculated at birth and average mass quantified (n = at least 4 mice per group).
Mice were weighed weekly, and growth is plotted as the average mass of each genotype/week (left: male, right: female, n > 5 per gender and genotype).
Mice were held by the tails and analyzed for the leg clasping phenotype. Representative images of Ccnd1+/+ and Ccnd1KI/KI age-matched mice are shown (n > 10 for each group).
Top: Individual whole-cell lysates were generated from the eyes of mice of each genotype and analyzed for expression of +/+ cyclin D1 (cyclin D1a) and KI/KI cyclin D1b via immunoblot using antisera specific to each isoform. Bottom: Representative H&E staining of retinal tissue from Ccnd1+/+ and Ccnd1KI/KI mice showing individual layers of the retina. The ganglion cell layer and inner plexiform layer (G + IPL), inner nuclear layer (INL), and outer nuclear layer (ONL) were quantified (right). Images were taken at 400× magnification.
Comparison of the phenotypes described in the Ccnd1−/− mouse with those observed in the Ccnd1KI/KI mouse, previously described (Sicinski et al, 1995).
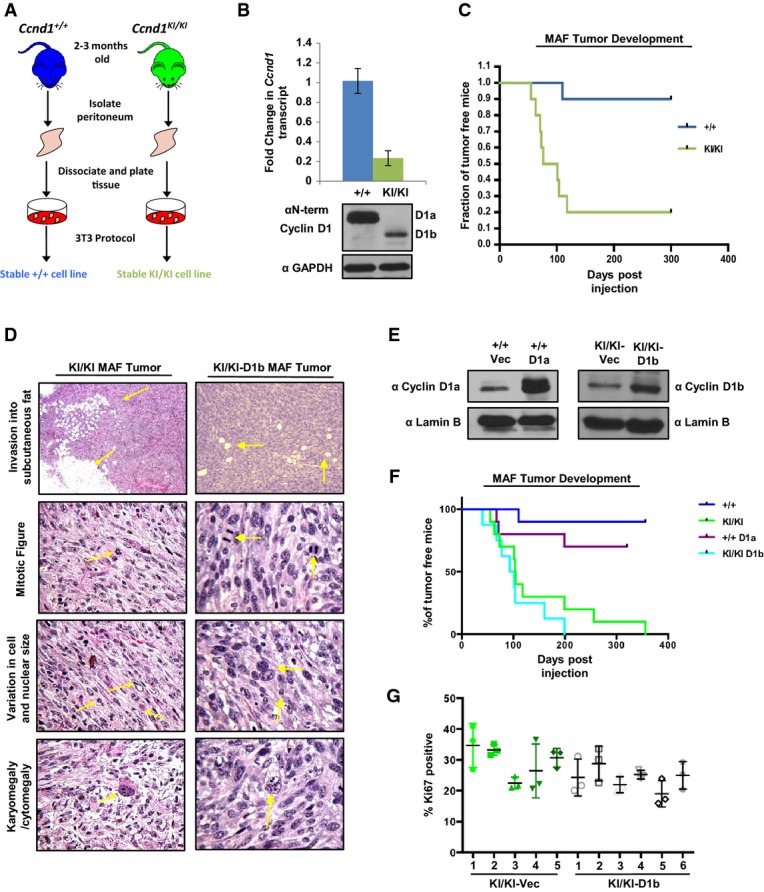
Murine adult fibroblasts (MAFs) were harvested from the peritoneum of Ccnd1+/+ and Ccnd1KI/KI mice, and stable cell lines were generated utilizing a 3T3 protocol.
Indicated passage-matched MAF lines were grown to 70% confluency and then harvested for RNA and protein extraction. Top: qPCR analysis of cyclin D1 transcript using primer pairs common to both transcript a and transcript b. Bottom: Immunoblot of cyclin D1 levels in Ccnd1+/+ and Ccnd1KI/KMAF lines using an antibody common to both isoforms.
Percentage of tumor-free mice post-injection of 1 million cells mixed with Matrigel (1:1 ratio) into the flanks of nude mice over a period of 10 months (n = 10 per genotype).
Indicated tumors were harvested 3 weeks after detectable tumor formation, fixed in formalin, and stained with H&E. Tumors were analyzed for features of malignancy by a board-certified veterinary pathologist. Arrows indicate the specific tumor-associated features noted. Top panels were taken at 40× magnification, and lower panels were taken at 400× magnification.
Ccnd1+/+ or Ccnd1KI/KI MAF lines were stably transfected with either cyclin D1a (+/+), cyclin D1b (KI/KI) or vector control constructs. After selection with puromycin, individual lines were assessed for the induction of cyclin D1a in the +/+ line and cyclin D1b in KI/KI lines via immunoblot.
One million cells from the indicated stable cell lines were injected subcutaneously into the flanks of nude mice and monitored for tumor formation over a period of 365 days. Indicated time points represent time of palpable tumor detection and are plotted as % of tumor-free mice over time.
Sections from KI/KI vec and KI/KI-D1b tumors were stained for the proliferative marker Ki67. Three random fields from each tumor were quantified for Ki67 positivity, and are plotted as the mean ratio of Ki67-positive cells/total cell number for each individual tumor.
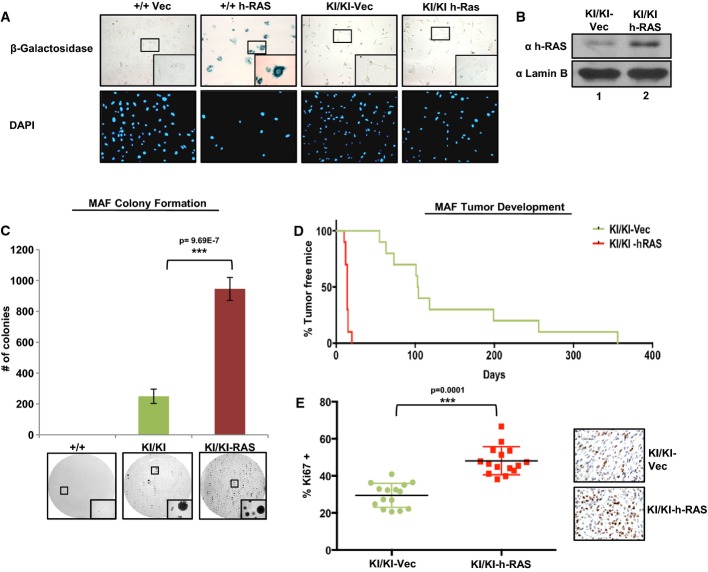
Passage-matched Ccnd1+/+ or Ccnd1KI/KI MAF lines were infected with lentivirus containing vector control or h-Ras constructs. Cells were selected with puromycin for 14 days and assayed for β-galactosidase activity. DAPI serves as a control for cell number. Images were taken at 200× magnification with insets taken at 400× magnification.
The indicated cell lines were grown in complete media and assayed for h-Ras expression via immunoblot. Lamin B serves as a loading control.
Cells were plated in soft agar and allowed to grow for a period of 3 weeks. Plates were then washed, fixed, and stained with 0.01% crystal violet. Colonies greater than 50 μm were counted and are plotted (top). Representative images of colony growth are shown for each cell line. Images were taken at 40× with insets taken at 200× magnification.
Indicated MAF lines were injected subcutaneously into the flanks of nude mice as in (A), and tumor incidence is plotted as % of tumor-free mice over time (n = 10).
Indicated tumors were harvested, fixed, and stained for the proliferative marker mKi67. Three random fields from each tumor were counted for mKi67 positivity, and are plotted as a % of total cell number, n = 5 (right: representative images). Images were taken at 400× magnification.
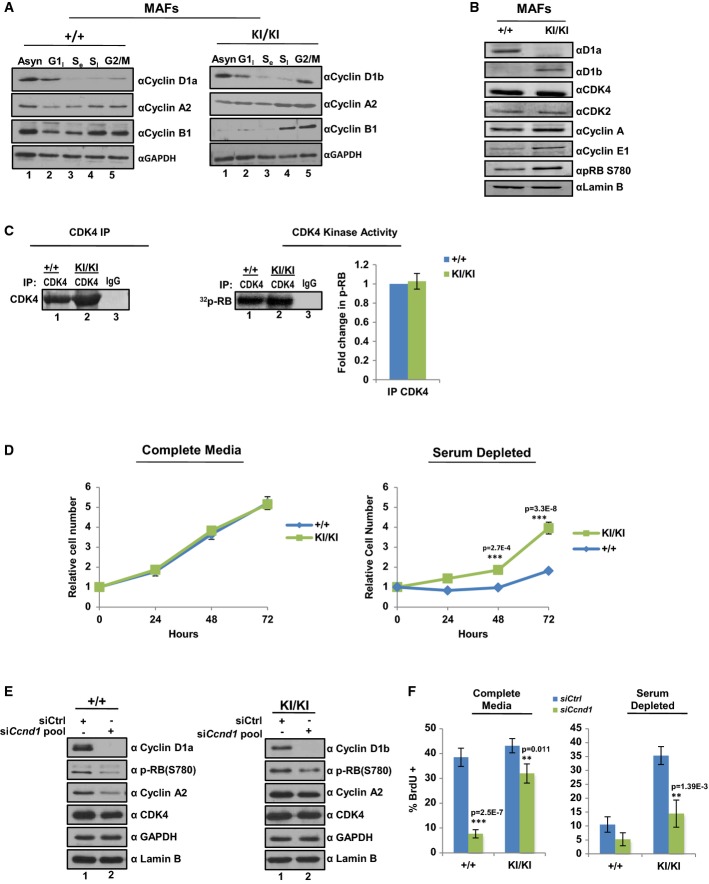
Ccnd1+/+ and Ccnd1KI/KIMAF lines were arrested in G1, S, and G2/M phases of the cell cycle and expression of cyclin D1 isoforms determined by immunoblot. cyclin A2 and cyclin B1 serve as phase-specific cell cycle controls.
Cells were plated in serum-proficient media and protein lysates generated from each genetic line. Individual D1 cyclins were immunoblotted to verify genetic identity along with other essential cell cycle components. Lamin B serves as a control.
Indicated cells lines were immunoprecipitated for CDK4 and analyzed for their ability to incorporate 32P-ATP into full-length RB substrate. Radioactive counts were normalized to CDK4 activity of +/+ cell lines and adjusted for efficiency of CDK4 pull down as determined by densitometry. Data is representative of three independent biological replicates.
Indicated cell lines were plated in serum-proficient (10%) or serum-deficient (1%) media and total cell number counted at 24, 48, and 72 h in biological triplicate.
Ccnd1+/+ and Ccnd1KI/KI MAF lines were transfected with a validated pool of siRNA directed against the N-terminus of the murine cyclin D1 transcript or control siRNA in full serum. 48 h post-transfection cells were harvested and analyzed for biochemical markers of cell cycle kinetics via immunoblot. GAPDH and Lamin B serve as controls.
Cells were treated as (E), and incubated in the indicated serum concentration for 48 h. Cells were treated with BrdU for 1 h prior to harvesting and then stained for BrdU incorporation. Three random fields from each of three biological triplicates were counted for BrdU incorporation via immunofluorescence and data is represented as percent positive/total cell number.
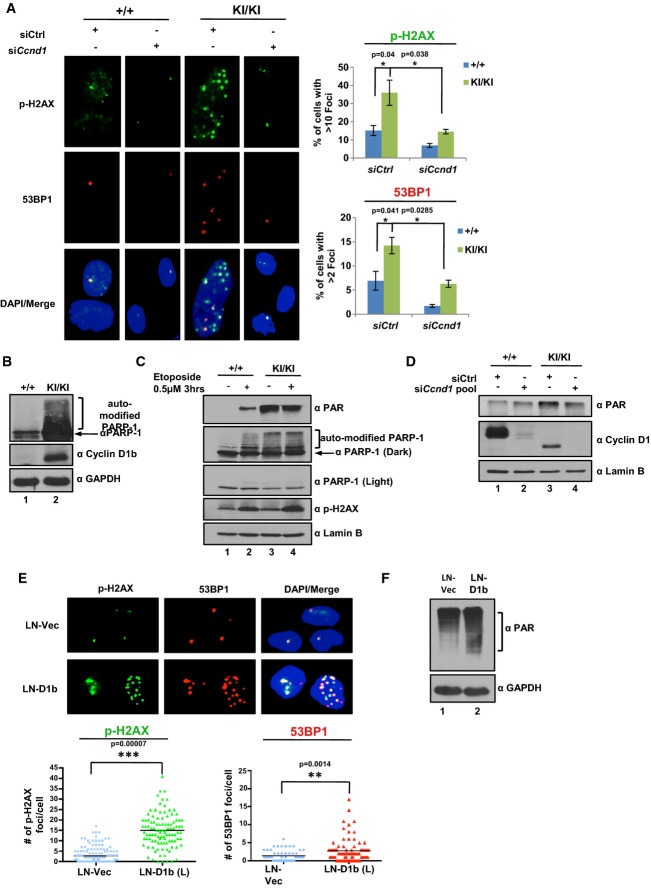
Ccnd1+/+ and Ccnd1KI/KI cells were transfected with a pool of siRNA directed against the N-terminus of the murine cyclin D1 transcript or scramble control for 48 h (as in Figure5). Cells were then fixed and stained for markers of double-strand breaks (p-H2AX and 53BP1, 400× objective). Total number of foci for each cell was quantified and represented as the % of cells with > 10 foci/cell (p-H2AX) or % of cells with > 2 foci/cell (53BP1). Error bars represent ± SEM.
Ccnd1+/+ and Ccnd1KI/KIMAF lines were grown in serum-proficient media for 24 h and probed for expression of auto-modified PARP1 via immunoblot. Gapdh and cyclin D1b serve as controls.
Cells were plated as in (B) and treated with 0.5 μM etoposide for 3 h. Cells were harvested and then probed for markers of DNA damage via immunoblot. Lamin B serves as a control.
Indicated MAF lines were treated as in (A) and harvested 48 h post-transfection. Cells were analyzed for total PAR levels via immunoblot. cyclin D1 levels serve as siRNA validation controls.
cyclin D1b expression was induced in the prostate cancer cell line LNCaP (previously described (Augello et al, 2014)) and stained for p-H2AX and 53BP1 foci as in (A) via immunofluorescence (400× objective). Total number of foci/cell is reported for LNCaP vector control and cyclin D1b-expressing isogenic pairs in biological triplicate.
Isogenic pairs from (E) were grown in serum-proficient media for 24 h, harvested, and probed for total PAR levels via immunoblot.
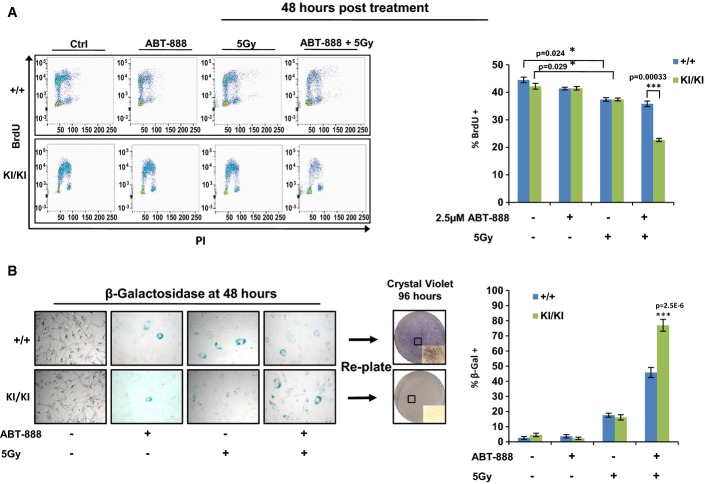
Ccnd1+/+ and Ccnd1KI/KIMAF lines were grown in serum-proficient media for 24 h. Cells were then treated with control DMSO or 2.5 μM of the PARP inhibitor ABT-888. One hour post-treatment, the indicated lines were treated with 5 Gy of radiation. Cells were then allowed to recover for 48 h, after which BrdU was added for 1 h, and then harvested for bivariate flow cytometry. Representative traces for each condition are shown (left) and BrdU incorporation of biological triplicates was quantified (right).
Cells were plated and treated as in (A). Forty-eight hours post-treatment, cells were fixed and stained for markers of senescence (β-galactosidase activity). Cells positive for the staining (blue) were quantified for each condition (400× magnification) and reported as a percentage of the total population (right). Plates treated in parallel with both IR and ABT-888 were harvested at 48 h via 1× trypsin and re-plated in serum-proficient media. Cells were allowed to grow for 96 h and were then stained with crystal violet (4× objective, with inset at 200× magnification, boxes highlight area of magnified images).
References
-
- Aggarwal P, Lessie MD, Lin DI, Pontano L, Gladden AB, Nuskey B, Goradia A, Wasik MA, Klein-Szanto AJ, Rustgi AK, et al. Nuclear accumulation of cyclin D1 during S phase inhibits Cul4-dependent Cdt1 proteolysis and triggers p53-dependent DNA rereplication. Genes Dev. 2007;21:2908–2922. - PMC - PubMed
-
- Arnold A, Papanikolaou A. Cyclin D1 in breast cancer pathogenesis. J Clin Oncol. 2005;23:4215–4224. - PubMed
-
- Benzeno S, Lu F, Guo M, Barbash O, Zhang F, Herman JG, Klein PS, Rustgi A, Diehl JA. Identification of mutations that disrupt phosphorylation-dependent nuclear export of cyclin D1. Oncogene. 2006;25:6291–6303. - PubMed
-
- Bertoni F, Rinaldi A, Zucca E, Cavalli F. Update on the molecular biology of mantle cell lymphoma. Hematol Oncol. 2006;24:22–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
