

Review
doi: 10.1021/cr500638n.
Epub 2015 Mar 19.
Amyloid β Protein and Alzheimer's Disease: When Computer Simulations Complement Experimental Studies
Affiliations
- PMID: 25789869
- PMCID: PMC5504919
- DOI: 10.1021/cr500638n
Item in Clipboard
Review
Amyloid β Protein and Alzheimer's Disease: When Computer Simulations Complement Experimental Studies
Chem Rev.
.
Abstract
Figures
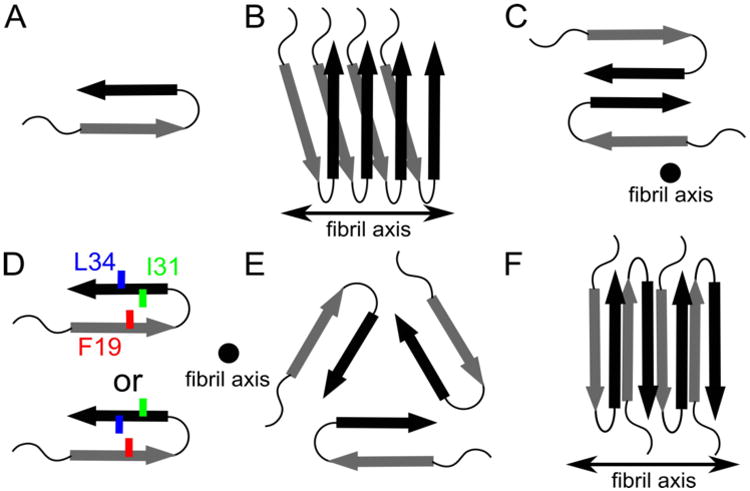
Schematics describing known structural motifs for Aβ1-40 WT and Aβ1-40 D23N fibrils. Arrows, thin lines, and colored symbols represent β-strand regions, non-β-strand regions, and selected residues, respectively. (A) Molecular conformation of Aβ molecules within fibrils, with arrows representing β-strand regions. (B) Organization of Aβ monomers into a protofilament. Each Aβ peptide contributes two β-strands to two stacked in-register parallel β-sheets, with hydrogen bonding interactions between equivalent β-strands along the fibril axis. (C) Cross-section of the Aβ1-40 fibril model of Petkova et al. composed of two protofilaments.(27,29) (D) Two distinct side chain arrangements experimentally observed for different Aβ1-40 fibrils. (E) Fibril cross-section predicted of the Aβ1-40 model determined by Paravastu et al. and composed of three protofilaments.(67) (F) The antiparallel β -sheet arrangement reported by Qiang et al. for fibrils of the Iowa Aβ1-40 D23N peptide.(29,101)
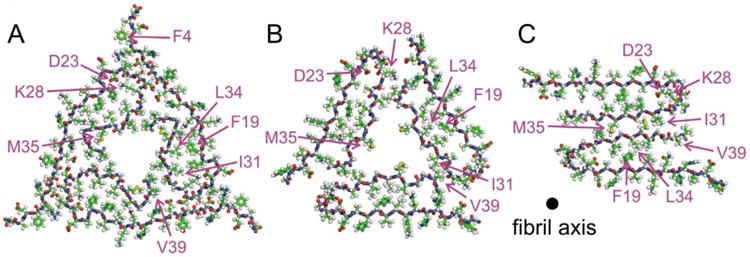
All-atom pictures of cross-sections of brain-derived Aβ1-40 fibrils modeled by Lu et al.(29) (A), compared to cross-sections of models for in vitro generated fibrils reported by Paravastu et al.(67) (B) and Petkova et al.(27) (C). The fibrils in (B) and (C) correspond to the diagrams shown in Figure 1C and 1E, respectively.
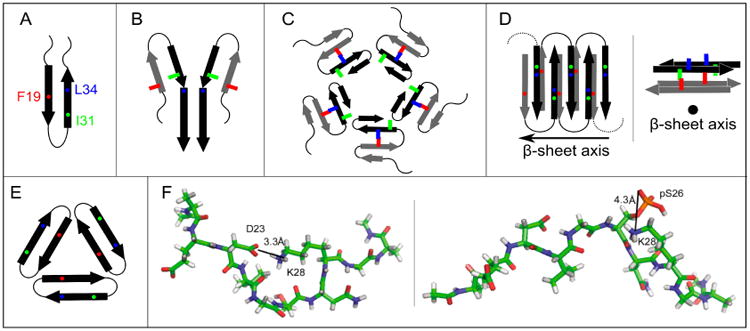
Proposed models for non-fibrillar Aβ aggregates. As in Figure 1, arrows, thin lines, and colored symbols represent β-strand and non-β-strand regions, and selected residues, respectively. (A) The antiparallel β-hairpin conformation predicted by Hoyer et al. for the monomer(114) and suggested by Scheidt et al. for protofibrils.(120,121) (B) The dimer structure proposed for preglobulomers by Yu et al.(123). (C) The disc-shaped pentamer model proposed by Ahmed et al.(124). (D) Two different views of the antiparallel β-sheet model for 150 kDa oligomers, reported by Tay et al.(111). (E) X-ray crystallographic structure of the trimer of the designed cyclic Aβ17-36 peptide.(127b) (F) Representative structures of highest populations in the MD ensembles of the Aβ21-30 WT peptide with the D23-K28 salt bridge (left) and the Aβ21-30 peptide with pS26 substitution (right).(47)
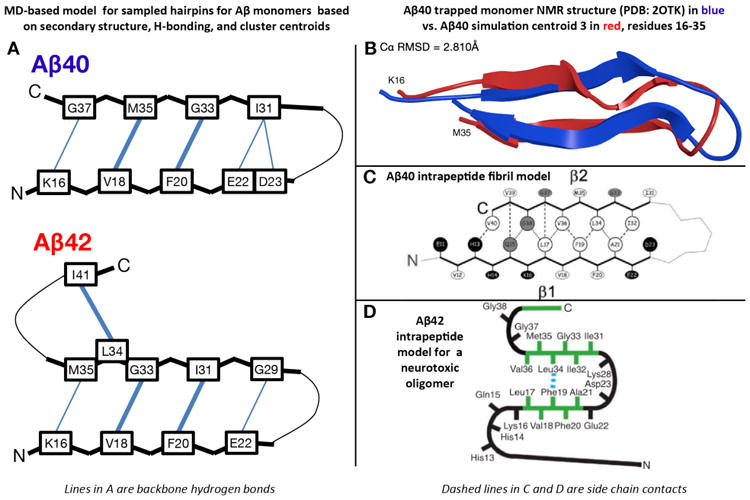
Transient REMD-sampled conformations of Aβ monomers bearing similarity to experimental intrapeptide models of higher order aggregates. (A) β-hairpin models for Aβ1-42 and Aβ1-40 monomers derived from the simulations of Rosenman et al.,(154b) based on the most populated cross-region backbone hydrogen bonds and secondary structure proclivities in the ensemble. Residues that have a high population for both donor to acceptor and acceptor to donor backbone hydrogen bonds are illustrated with a bold line. Sampled conformations matching these models exist as high-ranking centroid structures. (B) Ribbon overlay of residues 16-35 for centroid 3 derived from clustering analysis of the Aβ1-40 simulation(154b) and the solution NMR structure of monomeric Aβ1-40 in complex with a phage selected affibody (PDB entry 2OTK) published by Hoyer et al.(114). (C) Intrapeptide model for Aβ1-40 fibrils based on ss-NMR, as published by Bertini et al.(98). (D) Intrapeptide model for Aβ1-42 “on pathway” pentamers based on ss-NMR, as published by Ahmed et al.(124).
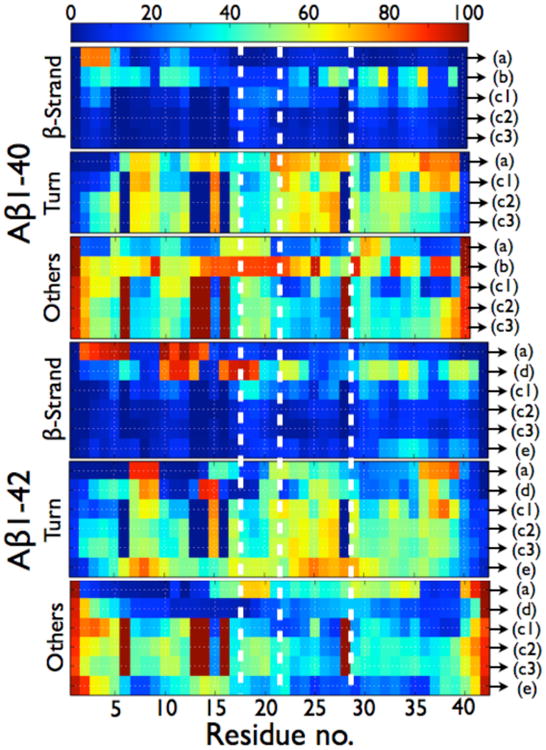
Per residue probabilities of β-strand, turn and others (coil and α-helix) for the Aβ1-40 and Aβ1-42 dimers as computed by different simulation protocols. As seen in Table 2, the total α-helix content amounts to less than 1.3% in Ref. 195, , and so others represent coil. In ref. 195, there is a total of 4.4% of α-helix in the dimers of Aβ1-42 and there is a probability of 27% for residues 23-28 to form α-helix. In Ref. 204, there is a total of 8.4% of α-helix in the dimers of Aβ1-42 and there is a probability of 45% for residues 13-17 to form a α-helix. (A) HT-REMD with the CG OPEP model and implicit solvent.(195) (B) united-atom CHARMM19 REMD with the SASA implicit solvent.(199) (C1) CG DMD with implicit solvent.(203) (C2) all-atom MD with OPLS-AA/SPCE.(204) (C3) all-atom MD with OPLS-AA/TIP3P.(204) (D) All-atom MC with implicit solvent.(181a) (E) all-atom REMD with OPLS-AA/TIP3P.(207) All secondary structure probabilities were computed using STRIDE; except for (b) for which the secondary structures are determined using information on the φ and Ψ dihedral angles only, without consideration of the H-bond network. As such, the β-sheet probabilities for (b) must be considered as extended conformations and no turn probability can be determined. The vertical dotted white lines delimit the four regions: N-terminal (residues 1-16), CHC (residues 17-21), loop region in the fibril (residues 22-28), and C-terminal (residues 29-40/42).
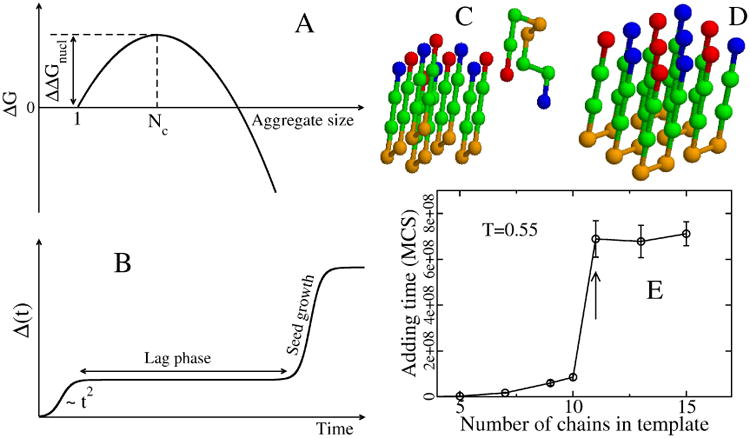
(A) Schematic plot of free energy of the aggregate, relative to the monomer, as a function of the aggregate size. The critical nucleus size corresponds to the peak of ΔG, while ΔΔGnucl is the barrier to nucleation. (B) Time dependence of the fibril mass M(t). Within homogeneous nucleation theory M(t) ∼ t2 at short time scales. The plateau corresponds to the lag phase, whose duration is proportional to exp(ΔΔGnucl/kBT). (C) A typical initial conformation for the (5+1) system in the lattice models with 8-bead sequence +HHPPHH-, where + and − refer to charged residues, while H and P denote hydrophobic and polar residues.(229) (D) The final fibril conformation with the lowest energy. (E) Dependence of the adding time τadd on the number of monomers that belong to the preformed template. Results are averaged over 50 Monte Carlo trajectories. At a concentration of 290 μM, τadd becomes independent of N for Ntemplate larger than 11. Arrow refers to Nc=11.(230)
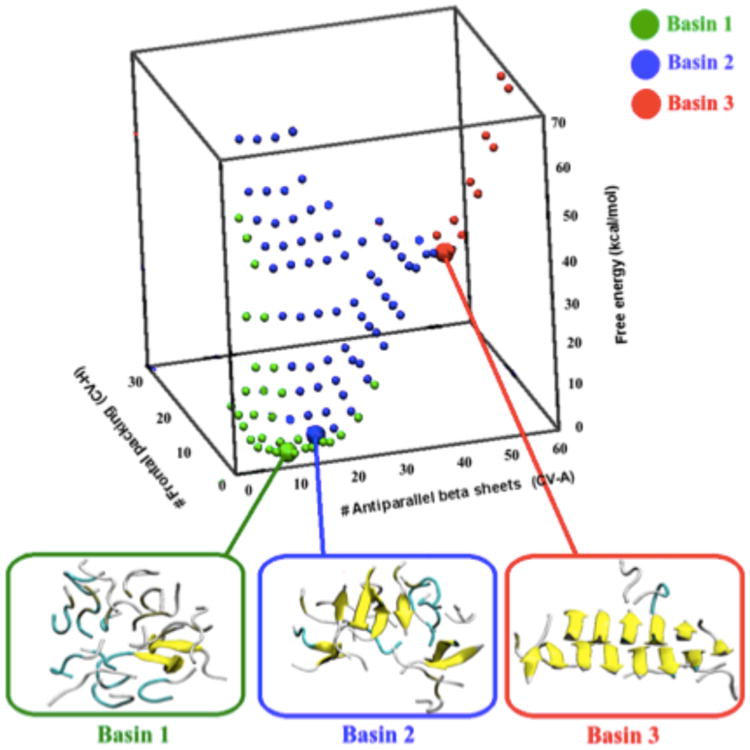
Free energy landscape of 18 Aβ35-40 peptides estimated by atomistic metadynamics simulations with explicit solvent as a function of the number of antiparallel β-sheets and the number of number of antiparallel frontal packing (different β-sheets facing each other).(178a) Basin 1 includes structures that are mainly disordered, with secondary structure elements formed only transiently. Basin 2 contains a much larger fraction of antiparallel β-sheets (up to 10–12 β-strands). The main characteristic of this basin is that the β-strands, although common, are not organized in a stable nucleus. These structures are only metastable, and can convert to the disordered melt in a few tenths of ns. Basin 3 includes instead structures with a specific inter-digitation of the side chains, which are stable at least on the ms time scale.
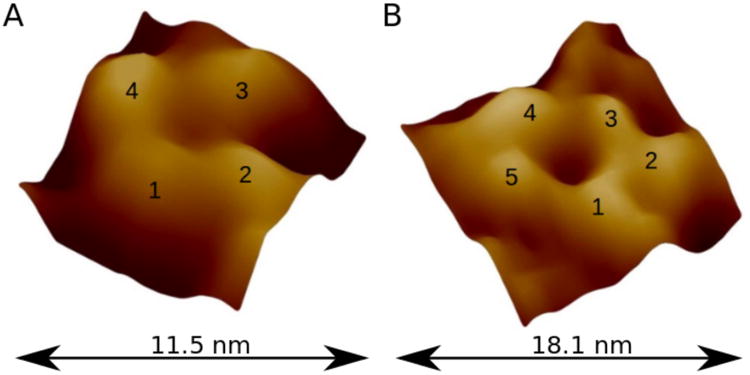
High-resolution AFM images of individual Aβ1-42 channels. They are most often observed as (A) tetrameric, or (B) pentameric subunits assemblies. Other types (e.g., hexameric) of pore-like Aβ structures were also reported.(269)
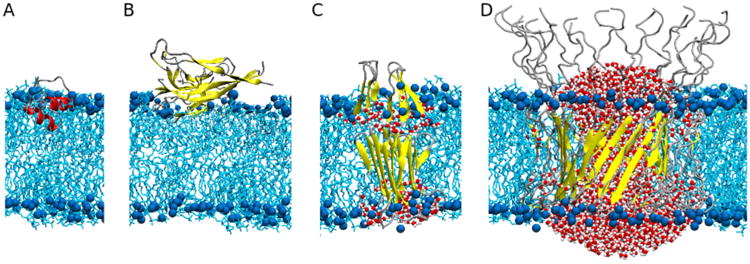
Various models of membrane-bound Aβ studied by MD simulations. (A) Monomeric Aβ1-40 is localized as a helical structure at the interface between membrane and solvent with the C-terminal helix inserting into the membrane.(288) (B) A fibrillar Aβ1-40 oligomer interacting with the membrane on the surface causes both a loss of β structure and a thinning of the membrane.(273) (C) The membrane-inserted β -sheet tetramer composed of Aβ1-42 allows water to permeate through the membrane(297) and can further assemble into a pore structure.(296) (D) Aβ1-42 barrel structure leading to pore formation in the membrane, which allows water and ion transport across the membrane.(293,294) Note that structures shown here are simulation-based models, as experimental high-resolution structures of membrane-bound Aβ peptides have yet to be resolved. The structural and mechanistic relationships between the simulation models are unknown. Aβ peptides are represented according to their local secondary structure: helices (red), β-sheets (yellow), and turns (silver). The bilayer phosphorus atoms (blue) are shown as van der Waals spheres and the lipid tails as licorice (cyan). For clarity, only water molecules inside the membrane are shown (van der Waals spheres).
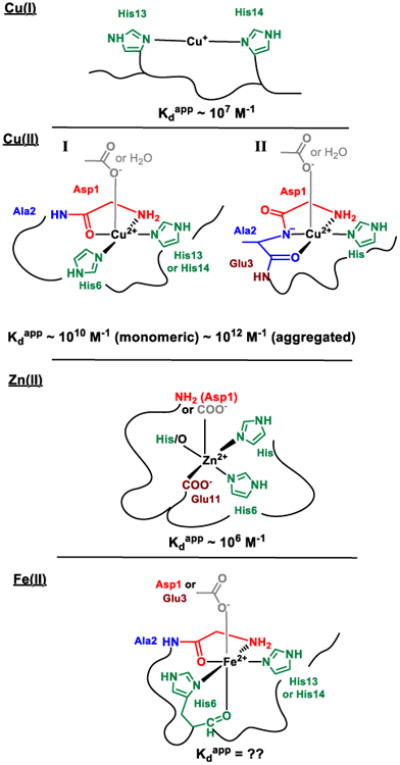
Models of the coordination sphere of different monomeric metal-Aβ complexes. Kdapp stands for apparent dissociation constant (i.e. dissociation constant at pH 7.4 in 0.1M salt and in the absence of buffer)
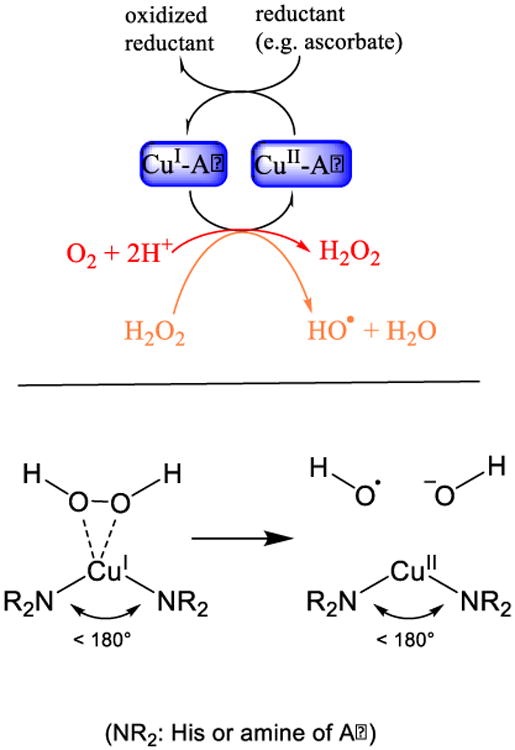
Catalytic role of Cu-Aβ in the production of reactive oxygen species. Top: Cu-Aβ is able to catalyze the production of H2O2 and HO° in the presence of a reducing agent and dioxygen. Bottom: The catalytic active state capable to catalyze the Fenton type reaction. Active are only the conformations with a diagonal ligation by the peptide remote from a angle of 180° and in which the H2O2 binds to Cu(I) side on.
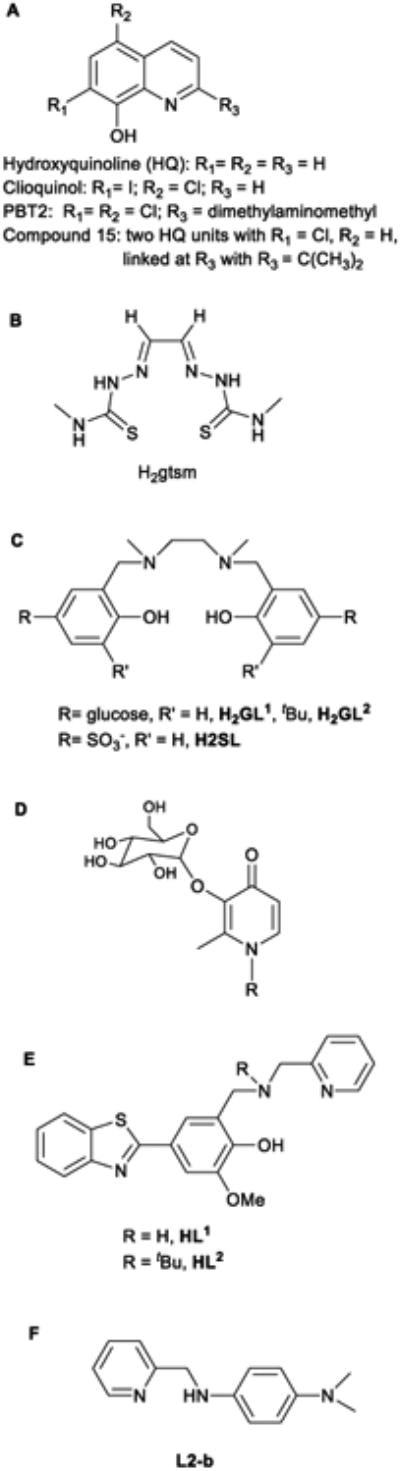
A selection of ligands studied in the context of metalmismetabolism in AD. A and B have ionophoric properties, a dimeric form(compound 15) is derived from clioquinol where the covalent attachment oftwo hydroxyquinoline increase Cu(II) affinity and selectivity, C are water-soluble Cu(II) chelators, C and D are brain-penetrating Cu(II) ligands and Eand F are bi-functional Cu(II) chelators with Aβ-targeting unit (for more detailssee Refs. 342 and 344).
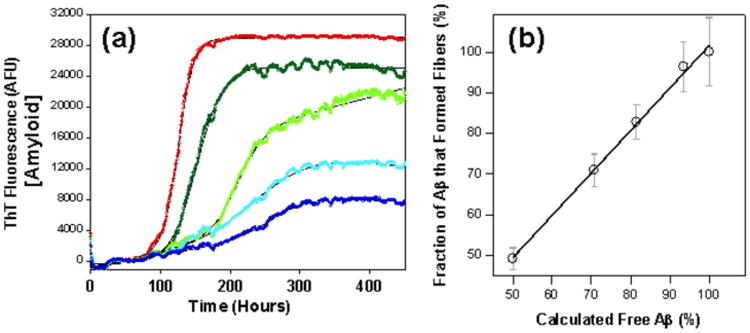
Physiological micro-molar concentrations of albumin inhibit fibre formation. (a) Kinetics of Aβ fibre formation in the presence of: no albumin (red); 1 μM (dark green); 3 μM (green); 5 μM (light blue); and 10 μM (dark blue) of albumin. (b) Competitive effects of albumin on total Aβ fibrils generated with a strong direct correlation between fraction of fibres generated and the calculated fraction of Aβ free to form fibres (not bound to increasing concentrations of albumin), based upon a Kd of 5 mM. Adapted from Stanyon et al.(352).
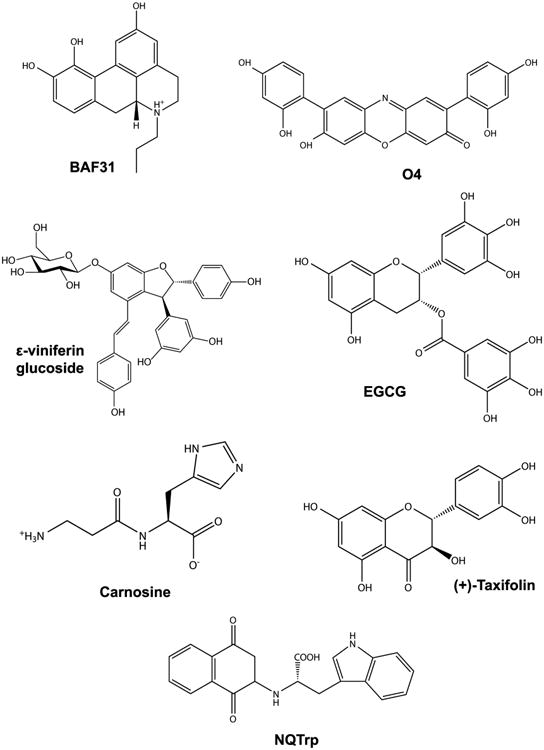
Chemical structures of small compound inhibitors of Aβ aggregation and toxicity.
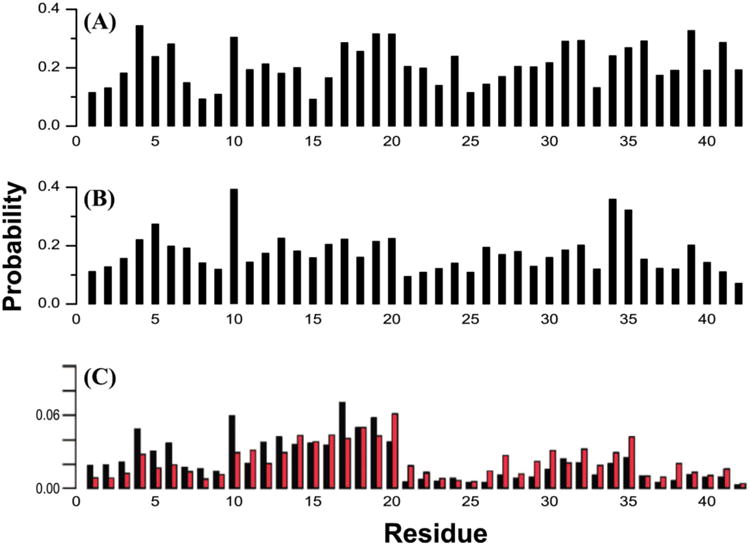
Contact probabilities of each Aβ1-42 each amino acid with compounds. (A) Between EGCG and Aβ1-42 dimer from atomistic REMD simulation.(207) (B) Between NQTrp and Aβ1-42 dimer from atomistic REMD simulation.(432) (C) Between 10 small organic molecular fragments (e.g., dimethyl ketone, furan and pyrazole) and Aβ1-42 monomer from REMD and FTMap.(175) Non-bonded (black bars) and hydrogen bonding (red bars) interactions are shown separately.
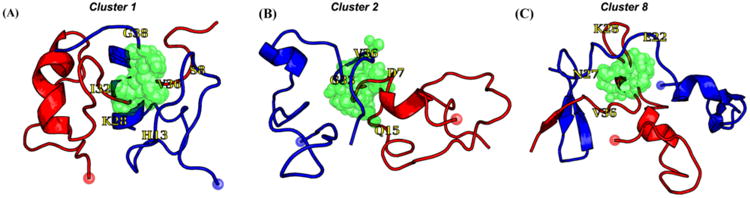
Representative structures of Aβ1-42 dimer/NQTrp from atomistic REMD simulations. (A) Cluster 1, (B) Cluster 2, and (C) Cluster 8. Chains A and B are colored red and blue, respectively. The red and blue spheres represent the Cα atoms of Asp1. The center of mass of NQTrp is shown as green spheres, and we show NQTrp if it forms a contact < 4 Å with any heavy atom of Aβ1-42. Residues colored yellow assist in the reading.
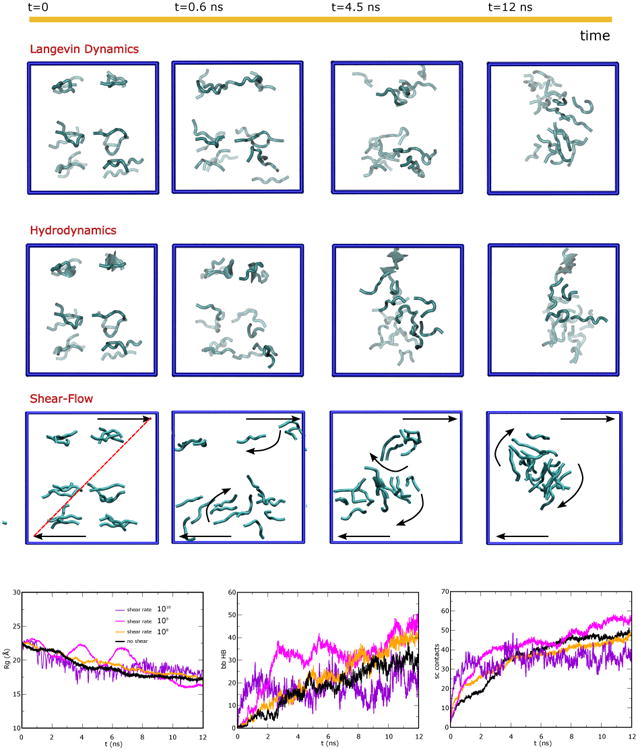
Aβ16-22 aggregation simulations under several conditions. Top panels. Pictorial representation of monomer aggregation for several simulation schemes: i) Langevin dynamics, ii) Langevin dynamics with Hydrodynamics Interactions (HI), and iii) Langevin dynamics with HI and longitudinal shear, shear rate Δv/Δz=108 s-1. The time-evolutions show the effect of HI on the aggregation kinetics. When shear-flow is activated also the details of the aggregation change: i.e. first two separate aggregates form and their relative motion in the shear flow (translation and rotation) guides their further encounter and fusion. Bottom panels. Time evolution of key parameters describing the aggregation process in shear-flow for different values of the shear rate Δv/Δz (0,108,109,1010 s-1): the gyration radius Rg (left panel), the number of inter-peptide hydrogen bonds formed between backbone NH and O groups (mid panel) and number of inter-peptide side-chain contacts (right panel). The system is composed of 18 peptides placed in a cubic box of L=65 Å and the shear gradient is generated along the Z direction.
References
-
- Alzheimer US Association. http://www.alz.org/
-
- Hardy J, Selkoe DJ. The Amyloid Hypothesis of Alzheimer's Disease: Progress and Problems on The Road To Therapeutics. Science. 2002;297:353–356. - PubMed
-
- Hernández-Rodríguez M, Correa-Basurto J, Martínez-Ramos F, Padilla-Martínez II, Benítez-Cardoza CG, Mera-Jiménez E, Rosales-Hernández MC. Design of Multi-target Compounds as AChE, BACE1, and amyloid-β(1-42) Oligomerization Inhibitors: in silico and in vitro Studies. J Alzheimers Dis. 2014;41:1073–1085. - PubMed
-
- Selkoe DJ. Alzheimer's Disease: Genes, Proteins, and Therapy. Physiol Rev. 2001;81:741–766. - PubMed
-
- Tan CC, Yu JT, Wang HF, Tan MS, Meng XF, Wang C, Jiang T, Zhu XC, Tan L. Efficacy and Safety of Donepezil, Galantamine, Rivastigmine, and Memantine for the Treatment of Alzheimer's Disease: A Systematic Review and Meta-analysis. J Alzheimers Dis. 2014;41:615–631. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
