

Epigenetic regulation of polyomavirus JC involves acetylation of specific lysine residues in NF-κB p65
- PMID: 25791343
- PMCID: PMC4575817
- DOI: 10.1007/s13365-015-0326-2
Epigenetic regulation of polyomavirus JC involves acetylation of specific lysine residues in NF-κB p65
Abstract
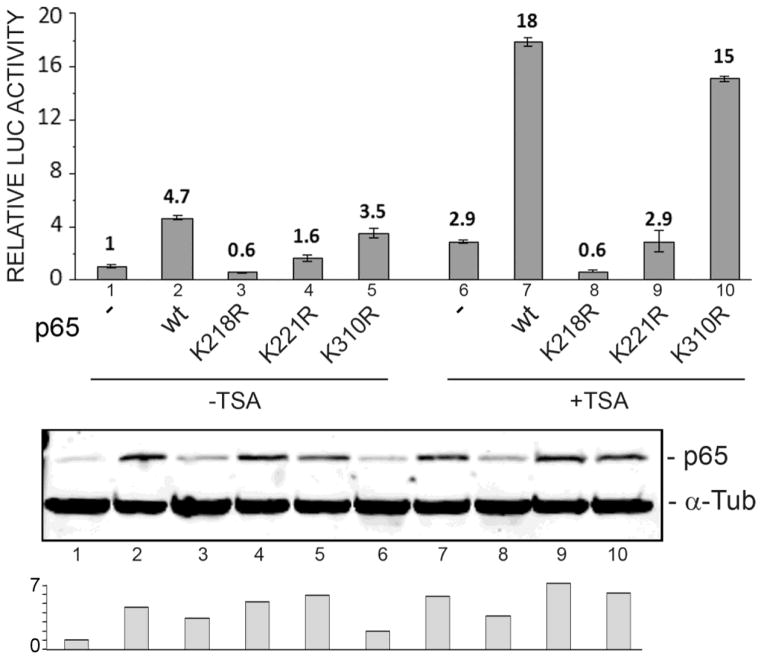
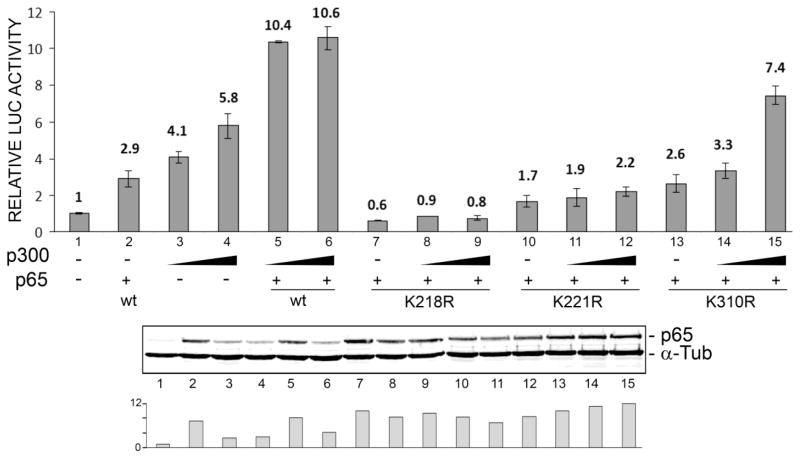
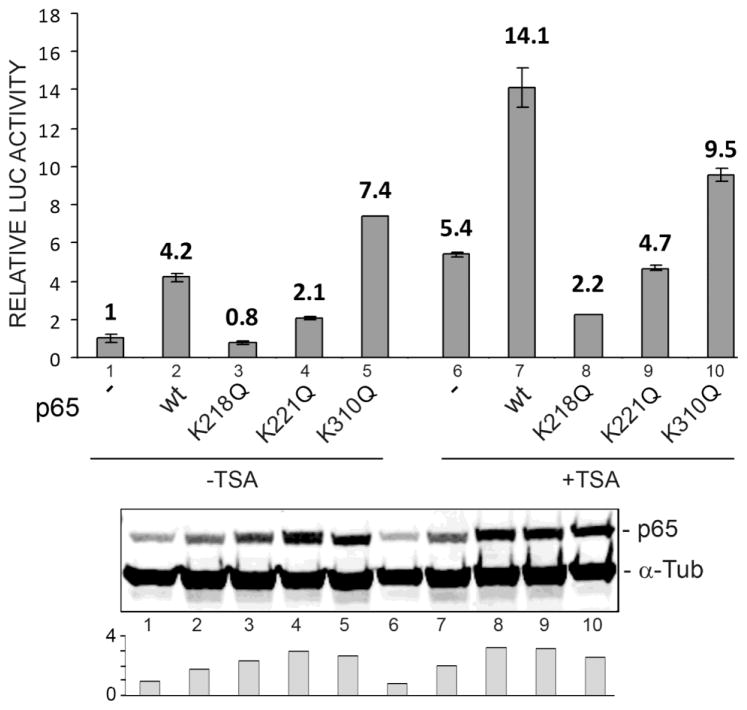
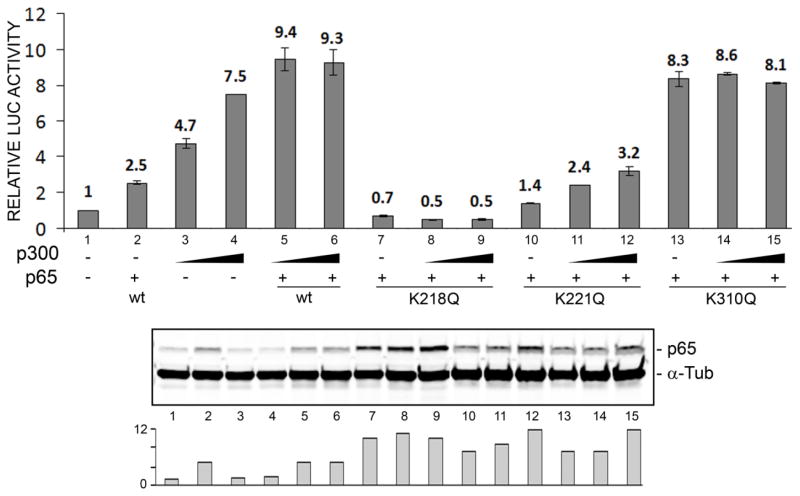
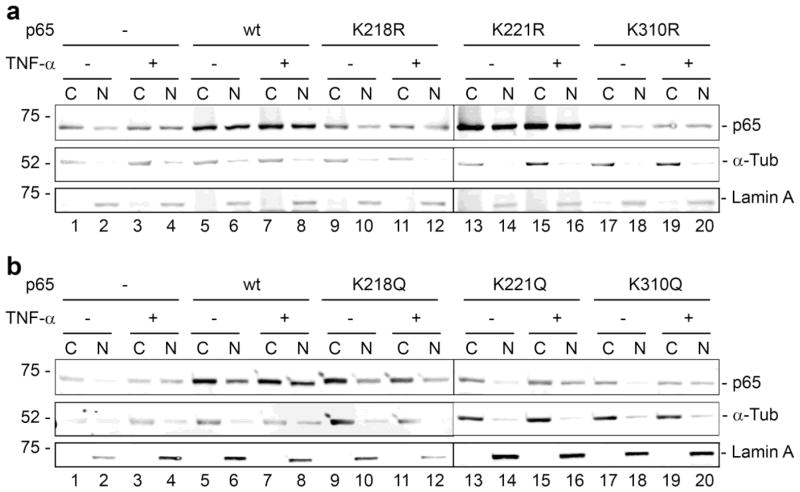
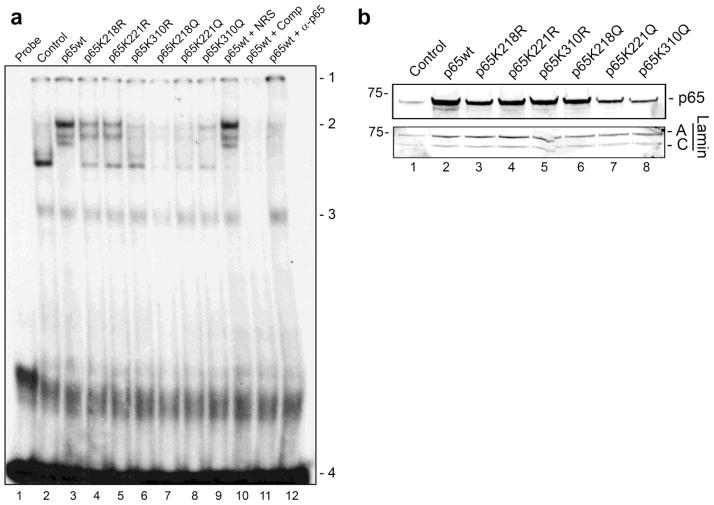
Progressive multifocal leukoencephalopathy (PML) is a fatal demyelinating disease caused by neurotropic polyomavirus, JC virus (JCV), a virus that causes lytic infection of CNS glial cells. After primary infection, JCV is controlled by the immune system but virus persists asymptomatically. Rarely, when immune function is impaired, it can reemerge to cause PML. The mechanisms of JCV persistence and reactivation are not well understood but our earlier work implicated epigenetic control by protein acetylation since histone deacetylase inhibitors such as trichostatin A (TSA) strongly stimulate JCV transcription. Since both TNF-α and TSA activate JCV transcription via the same unique NF-κB site in the JCV control region, we investigated a role for acetylation of NF-κB in JCV regulation. A site-directed mutagenesis strategy was employed targeting the known lysine acetylation sites of NF-κB p65: K218, K221, and K310. We individually mutated each lysine to arginine, which cannot be acetylated and retains a positive charge like lysine. K218R and K221R impaired transactivation of JCV early promoter transcription either alone or combined with TSA treatment or coexpression of acetyltransferase transcriptional coactivator p300 but K310R was largely without effect. Mutation of lysine to glutamine gives mutants with a negative charge like acetyllysine. However, K218Q and K221Q showed impaired activity and only K310Q showed enhanced transactivation. NF-κB acetylation can regulate several aspects of the process of activation including complex formation with IκB, translocation to the nucleus, and DNA binding and transcriptional activation. Cell fractionation studies revealed that the mutants had no defect in translocation to the nucleus whereas gel shift studies revealed reduced binding to the JCV NF-κB site. Thus, acetylation regulates NF-κB p65 activity toward JCV at the level of p65 binding to the JCV control region and activation of JCV transcription.
Keywords: JC Virus; Progressive multifocal leukoencephalopathy; Viral persistence; Viral reactivation.
Conflict of interest statement
CONFLICT OF INTEREST
The authors Hassen S. Wollebo, Anna Bellizzi, Dominique H. Cossari, Mahmut Safak, Kamel Khalili and Martyn K. White declare that they have no conflicts of interest.
Figures
References
-
- Berger JR. Progressive multifocal leukoencephalopathy and newer biological agents. Drug Saf. 2010;33:969–983. - PubMed
-
- Berger JR. The clinical features of PML. Cleve Clin J Med. 2011;78(Suppl 2):S8–12. - PubMed
-
- Calao M, Burny A, Quivy V, Dekoninck A, Van Lint C. A pervasive role of histone acetyltransferases and deacetylases in an NF-kappaB-signaling code. Trends Biochem Sci. 2008;33:339–349. - PubMed
-
- DeCaprio JA, Imperiale MJ, Major EO. Polyomaviruses. In: Knipe DM, Howley PM, editors. Fields Virology. 6. Lippincott, Williams & Wilkins; Philadelphia: 2013. pp. 6133–1661.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
