

Ethylene Signaling Influences Light-Regulated Development in Pea
- PMID: 25792252
- PMCID: PMC4577373
- DOI: 10.1104/pp.15.00164
Ethylene Signaling Influences Light-Regulated Development in Pea
Abstract
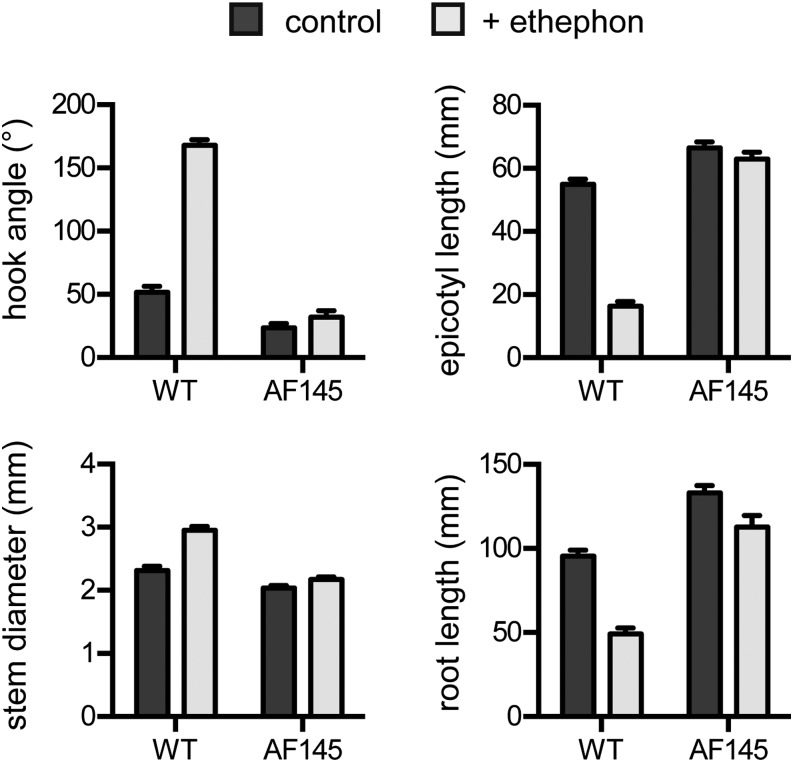
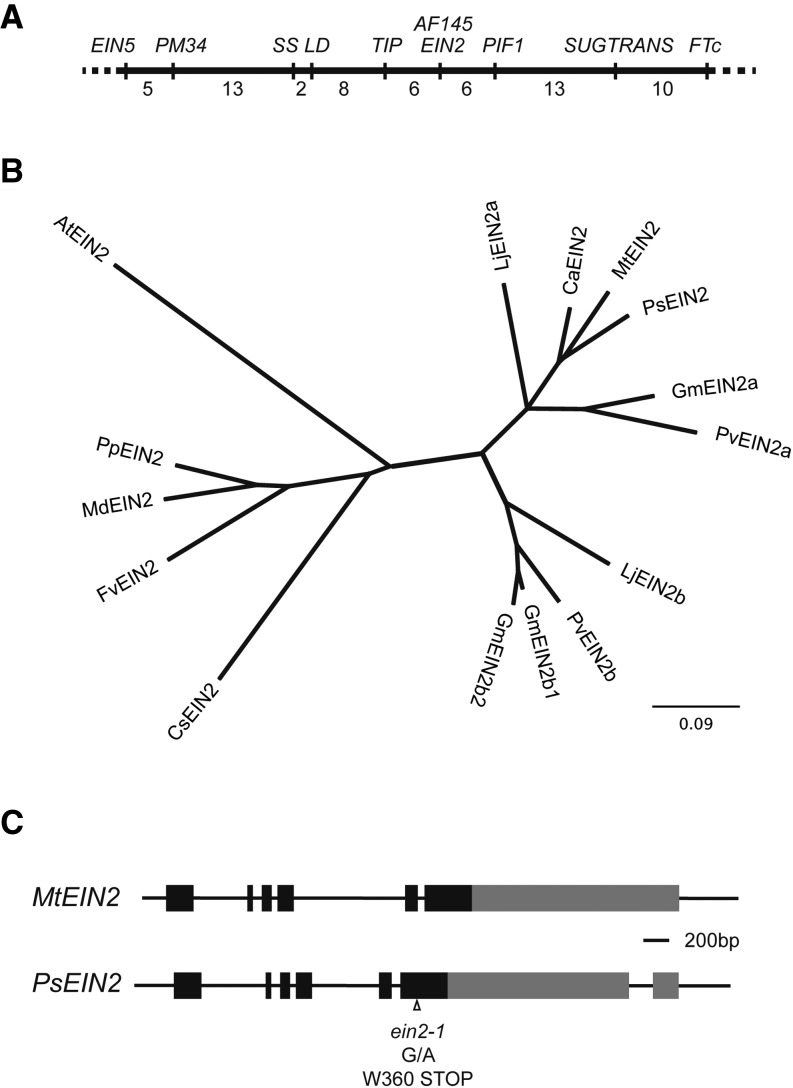
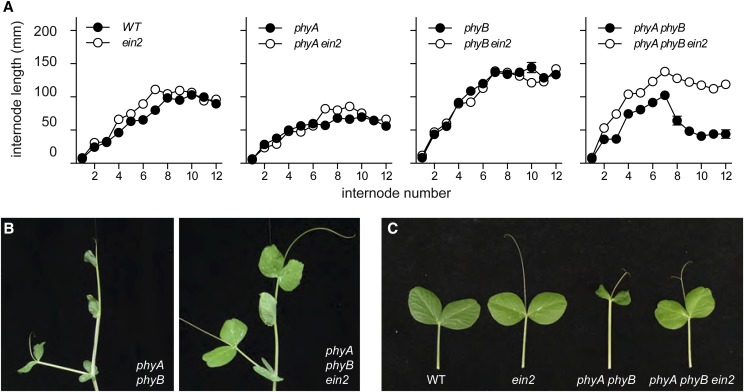
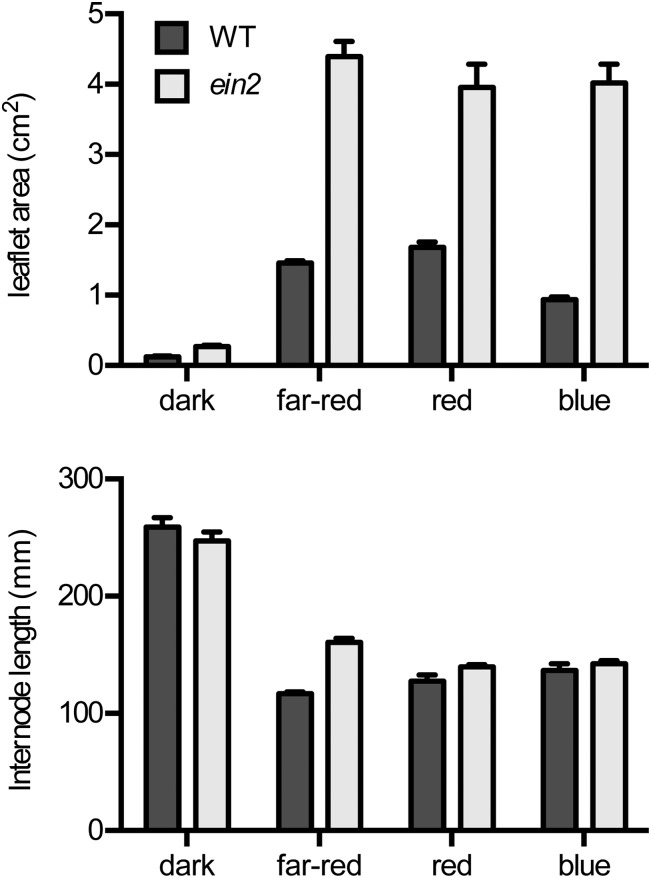
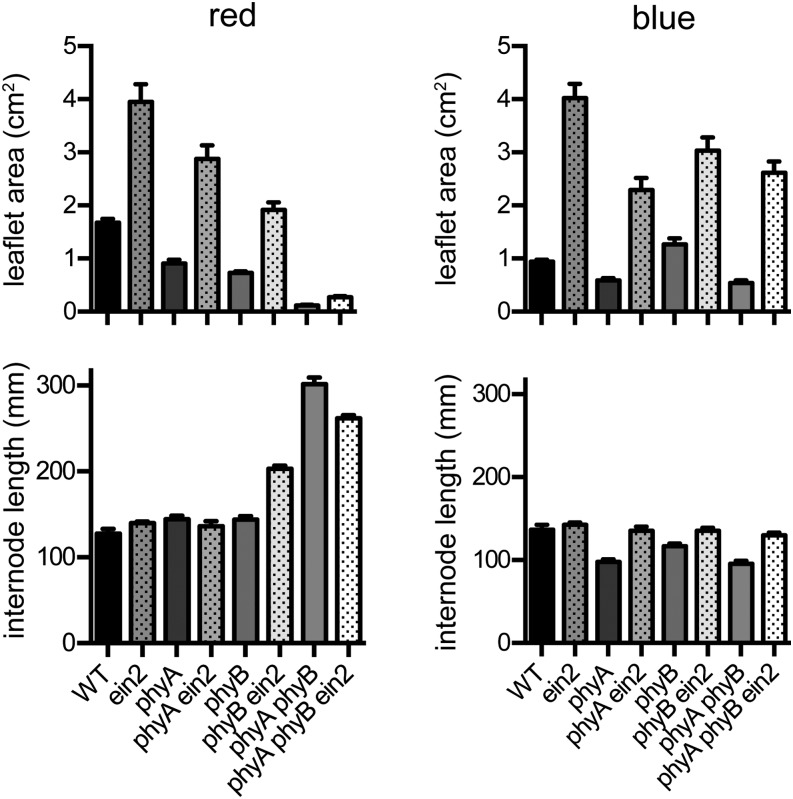
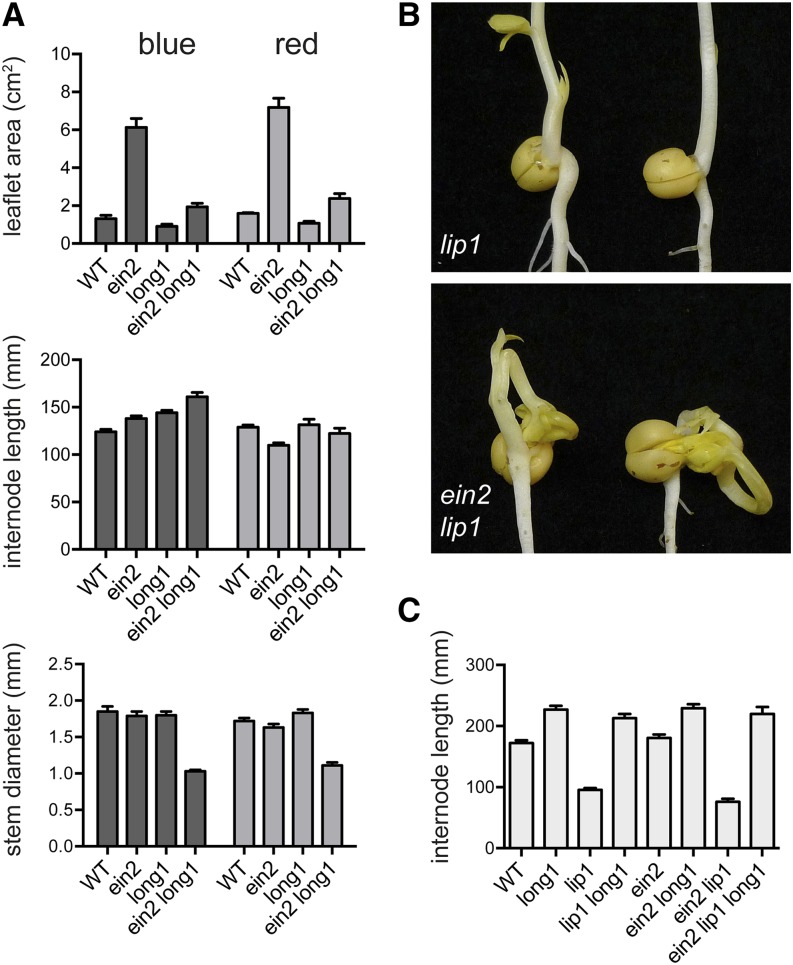
Plant responses to light involve a complex network of interactions among multiple plant hormones. In a screen for mutants showing altered photomorphogenesis under red light, we identified a mutant with dramatically enhanced leaf expansion and delayed petal senescence. We show that this mutant exhibits reduced sensitivity to ethylene and carries a nonsense mutation in the single pea (Pisum sativum) ortholog of the ethylene signaling gene ETHYLENE INSENSITIVE2 (EIN2). Consistent with this observation, the ein2 mutation rescues the previously described effects of ethylene overproduction in mature phytochrome-deficient plants. In seedlings, ein2 confers a marked increase in leaf expansion under monochromatic red, far-red, or blue light, and interaction with phytochromeA, phytochromeB, and long1 mutants confirms that ein2 enhances both phytochrome- and cryptochrome-dependent responses in a LONG1-dependent manner. In contrast, minimal effects of ein2 on seedling development in darkness or high-irradiance white light show that ethylene is not limiting for development under these conditions. These results indicate that ethylene signaling constrains leaf expansion during deetiolation in pea and provide further evidence that down-regulation of ethylene production may be an important component mechanism in the broader control of photomorphogenic development by phytochrome and cryptochrome.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures

References
-
- Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR (1999) EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284: 2148–2152 - PubMed
-
- Aubert G, Morin J, Jacquin F, Loridon K, Quillet MC, Petit A, Rameau C, Lejeune-Hénaut I, Huguet T, Burstin J (2006) Functional mapping in pea, as an aid to the candidate gene selection and for investigating synteny with the model legume Medicago truncatula. Theor Appl Genet 112: 1024–1041 - PubMed
-
- Bailey-Serres J, Fukao T, Gibbs DJ, Holdsworth MJ, Lee SC, Licausi F, Perata P, Voesenek LA, van Dongen JT (2012) Making sense of low oxygen sensing. Trends Plant Sci 17: 129–138 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
