

Suppression of sensitivity to drugs and antibiotics by high external cation concentrations in fission yeast
- PMID: 25793410
- PMCID: PMC4368599
- DOI: 10.1371/journal.pone.0119297
Suppression of sensitivity to drugs and antibiotics by high external cation concentrations in fission yeast
Abstract
Background: Potassium ion homeostasis plays an important role in regulating membrane potential and therefore resistance to cations, antibiotics and chemotherapeutic agents in Schizosaccharomyces pombe and other yeasts. However, the precise relationship between drug resistance in S. pombe and external potassium concentrations (particularly in its natural habitats) remains unclear. S. pombe can tolerate a wide range of external potassium concentrations which in turn affect plasma membrane polarization. We thus hypothesized that high external potassium concentrations suppress the sensitivity of this yeast to various drugs.
Methods: We have investigated the effect of external KCl concentrations on the sensitivity of S. pombe cells to a wide range of antibiotics, antimicrobial agents and chemotherapeutic drugs. We employed survival assays, immunoblotting and microscopy for these studies.
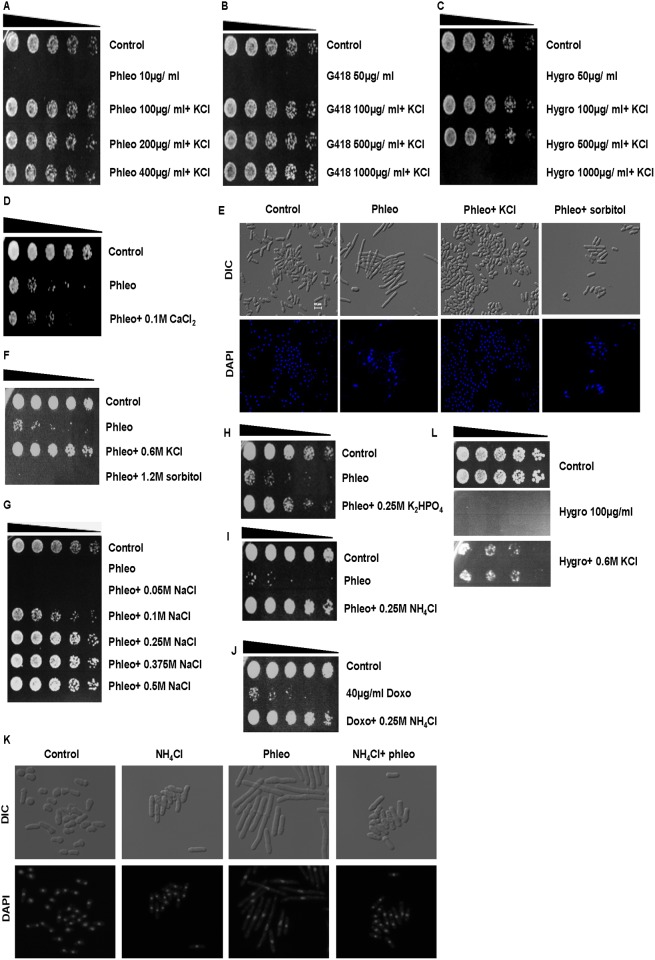
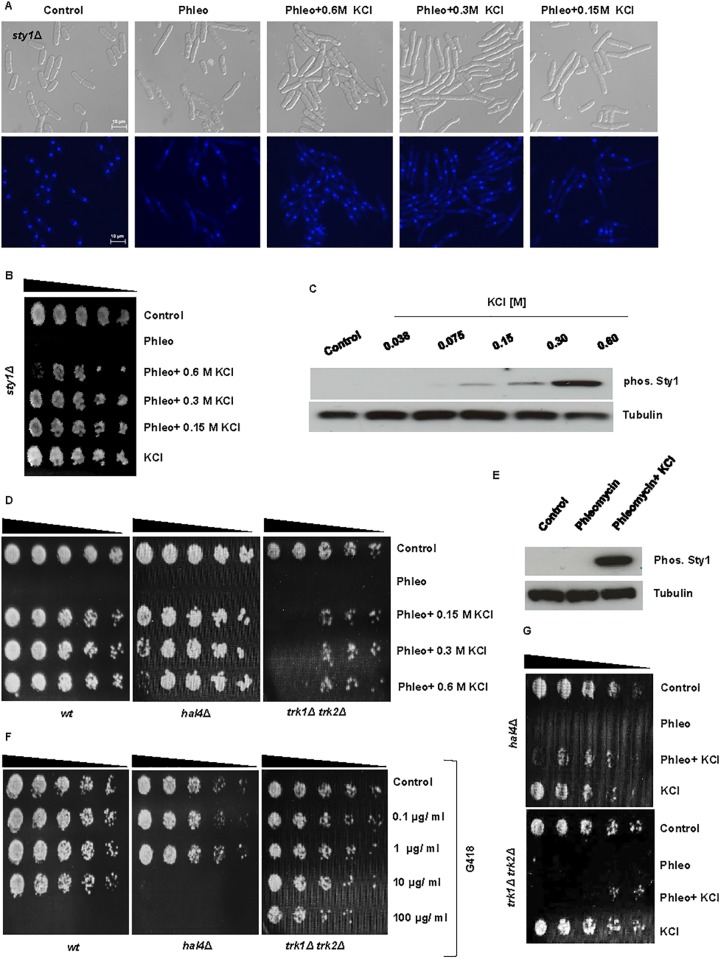
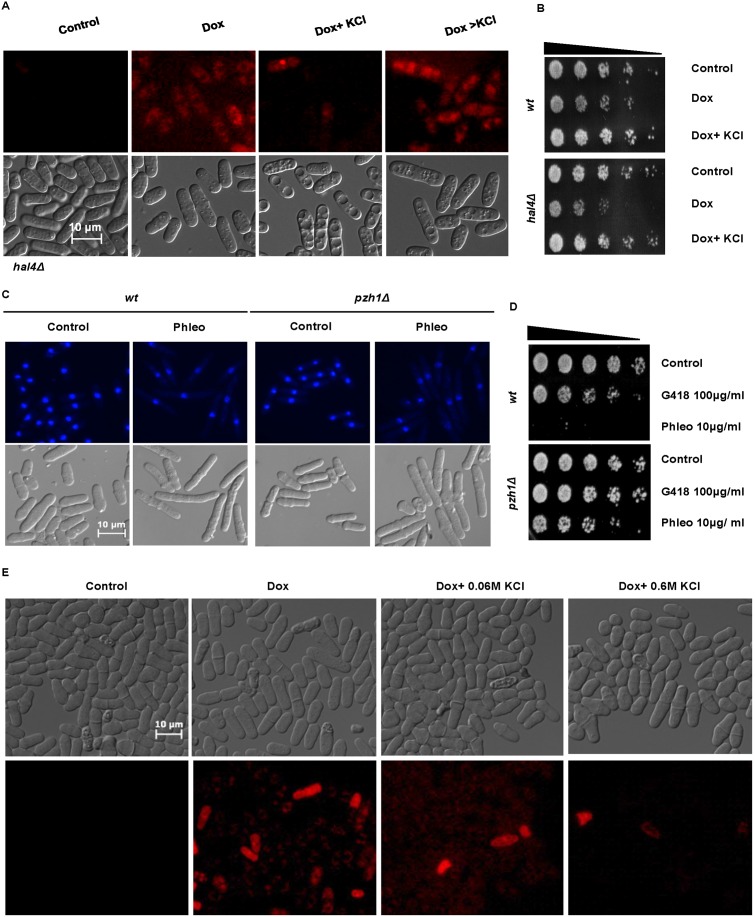
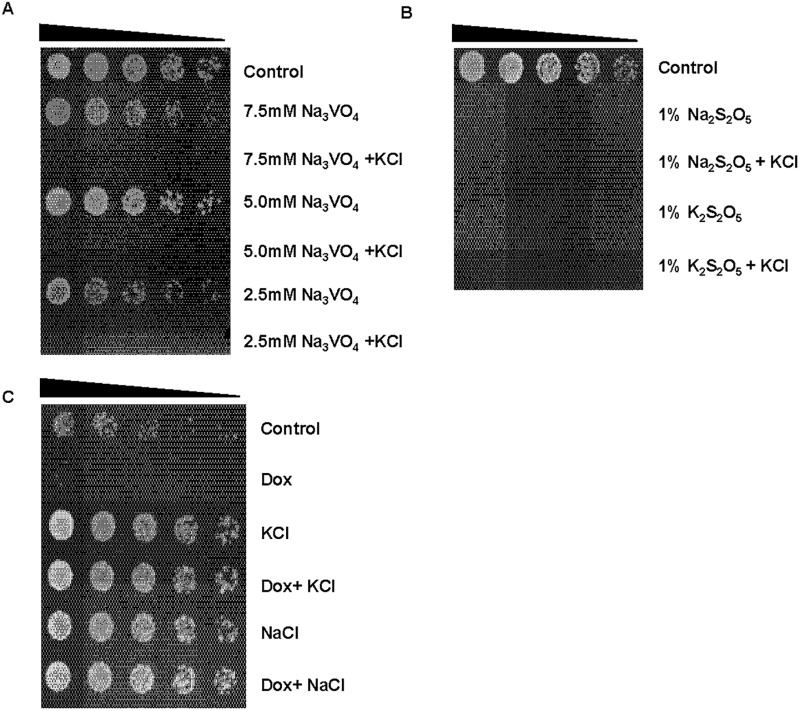
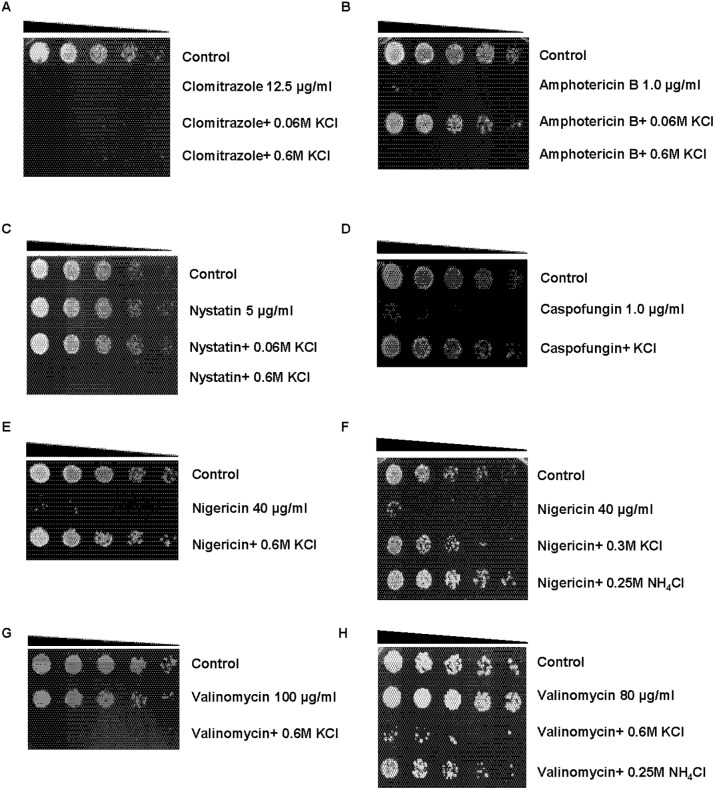
Results: We demonstrate that KCl, and to a lesser extent NaCl and RbCl can suppress the sensitivity of S. pombe to a wide range of antibiotics. Ammonium chloride and potassium hydrogen sulphate also suppressed drug sensitivity. This effect appears to depend in part on changes to membrane polarization and membrane transport proteins. Interestingly, we have found little relationship between the suppressive effect of KCl on sensitivity and the structure, polarity or solubility of the various compounds investigated.
Conclusions: High concentrations of external potassium and other cations suppress sensitivity to a wide range of drugs in S. pombe. Potassium-rich environments may thus provide S. pombe a competitive advantage in nature. Modulating potassium ion homeostasis may sensitize pathogenic fungi to antifungal agents.
Conflict of interest statement
Figures

References
-
- Aouida M, Tounekti O, Leduc A, Belhadj O, Mir L, Ramotar D. Isolation and characterization of Saccharomyces cerevisiae mutants with enhanced resistance to the anticancer drug bleomycin. Current genetics 2004, 45(5):265–272. - PubMed
-
- Bianchi L, Wible B, Arcangeli A, Taglialatela M, Morra F, Castaldo P, et al. herg encodes a K+ current highly conserved in tumors of different histogenesis: a selective advantage for cancer cells? Cancer research 1998, 58(4):815–822. - PubMed
-
- Lafon-Lafourcade S, Lonvaud-Funel A, Carre E. Lactic acid bacteria of wines: stimulation of growth and malolactic fermentation. Antonie van Leeuwenhoek 1983, 49(3):349–352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
