

Exceptionally widespread nanomachines composed of type IV pilins: the prokaryotic Swiss Army knives
- PMID: 25793961
- PMCID: PMC4471445
- DOI: 10.1093/femsre/fuu001
Exceptionally widespread nanomachines composed of type IV pilins: the prokaryotic Swiss Army knives
Abstract
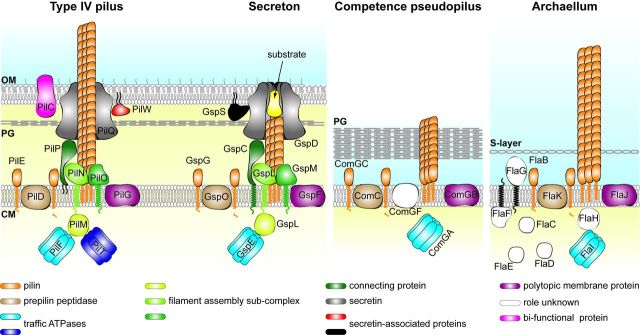
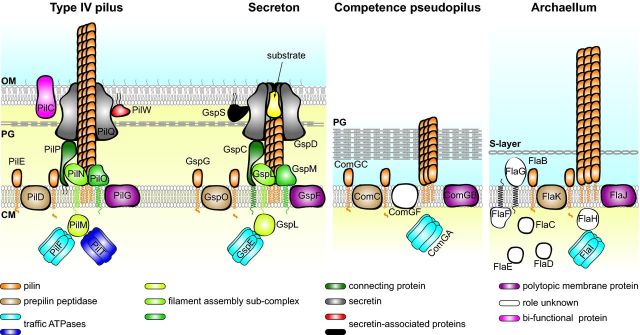
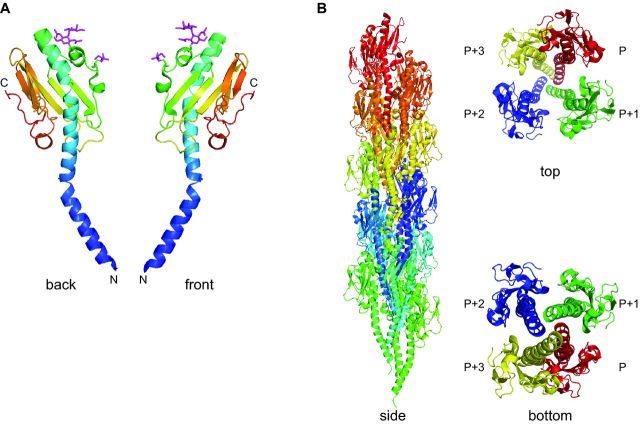
Prokaryotes have engineered sophisticated surface nanomachines that have allowed them to colonize Earth and thrive even in extreme environments. Filamentous machineries composed of type IV pilins, which are associated with an amazing array of properties ranging from motility to electric conductance, are arguably the most widespread since distinctive proteins dedicated to their biogenesis are found in most known species of prokaryotes. Several decades of investigations, starting with type IV pili and then a variety of related systems both in bacteria and archaea, have outlined common molecular and structural bases for these nanomachines. Using type IV pili as a paradigm, we will highlight in this review common aspects and key biological differences of this group of filamentous structures.
Keywords: archaellum; class III signal peptide; prepilin peptidase; type II secretion system; type IV pilus.
© The Author 2015. Published by Oxford University Press on behalf of FEMS.
Figures


References
-
- Aas FE, Winther-Larsen HC, Wolfgang M, et al. Substitutions in the N-terminal alpha helical spine of Neisseria gonorrhoeae pilin affect type IV pilus assembly, dynamics and associated functions. Mol Microbiol. 2007;63:69–85. - PubMed
-
- Abendroth J, Bagdasarian M, Sandkvist M, et al. The structure of the cytoplasmic domain of EpsL, an inner membrane component of the type II secretion system of Vibrio cholerae: an unusual member of the actin-like ATPase superfamily. J Mol Biol. 2004a;344:619–33. - PubMed
-
- Abendroth J, Murphy P, Sandkvist M, et al. The X-ray structure of the type II secretion system complex formed by the N-terminal domain of EpsE and the cytoplasmic domain of EpsL of Vibrio cholerae. J Mol Biol. 2005;348:845–55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
