

An O antigen capsule modulates bacterial pathogenesis in Shigella sonnei
- PMID: 25794007
- PMCID: PMC4368438
- DOI: 10.1371/journal.ppat.1004749
An O antigen capsule modulates bacterial pathogenesis in Shigella sonnei
Abstract
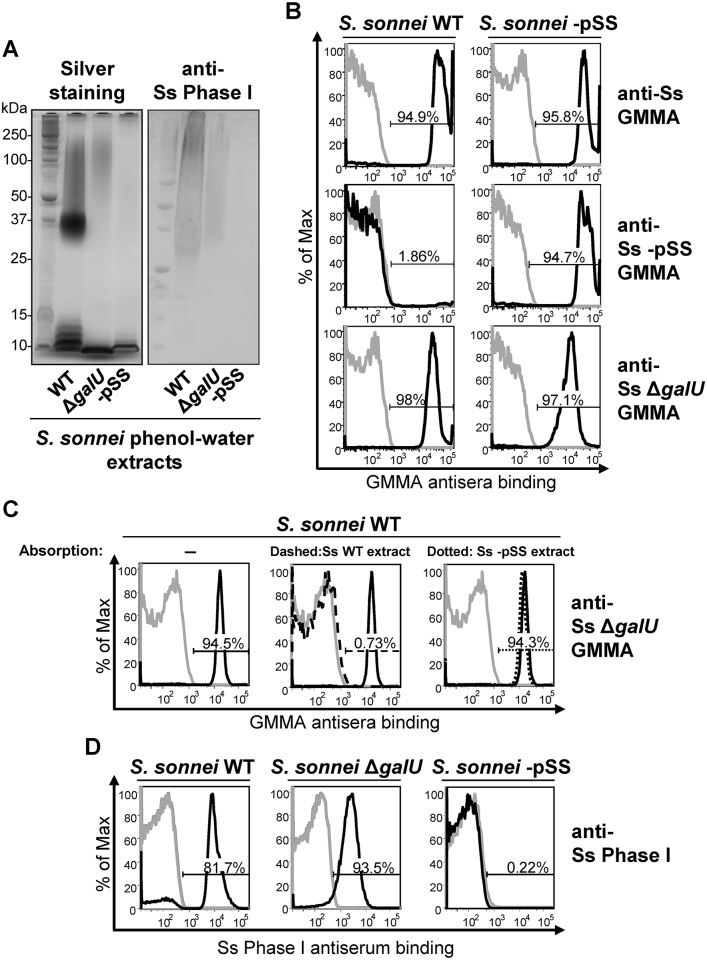
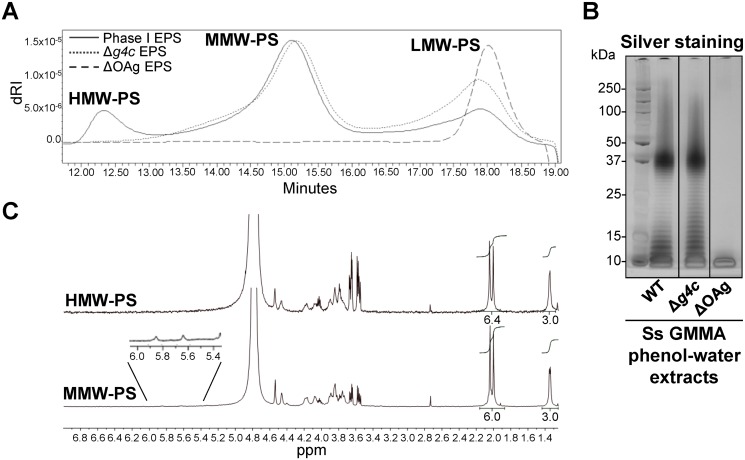
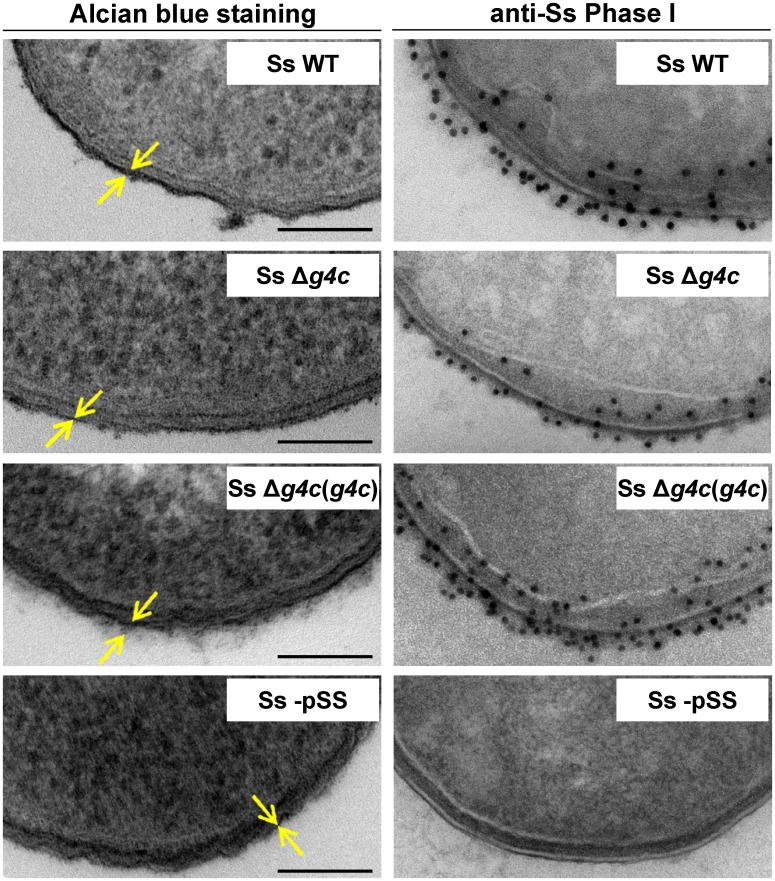
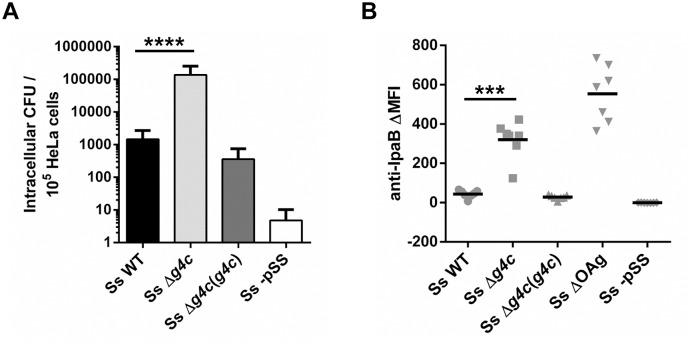
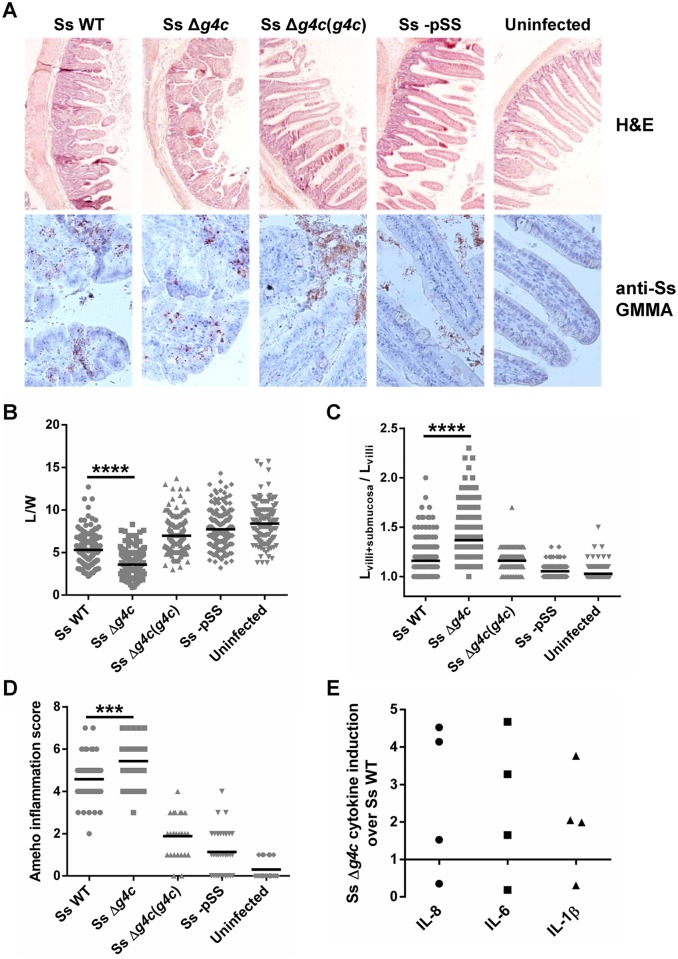
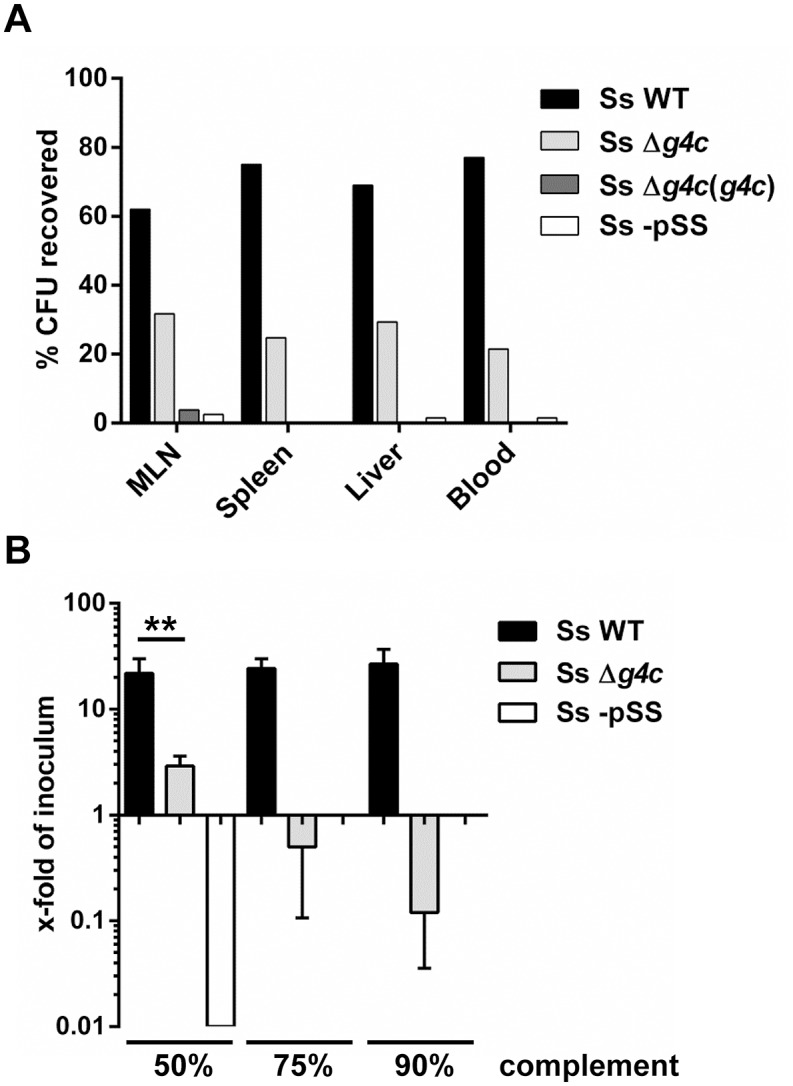
Shigella is the leading cause for dysentery worldwide. Together with several virulence factors employed for invasion, the presence and length of the O antigen (OAg) of the lipopolysaccharide (LPS) plays a key role in pathogenesis. S. flexneri 2a has a bimodal OAg chain length distribution regulated in a growth-dependent manner, whereas S. sonnei LPS comprises a monomodal OAg. Here we reveal that S. sonnei, but not S. flexneri 2a, possesses a high molecular weight, immunogenic group 4 capsule, characterized by structural similarity to LPS OAg. We found that a galU mutant of S. sonnei, that is unable to produce a complete LPS with OAg attached, can still assemble OAg material on the cell surface, but a galU mutant of S. flexneri 2a cannot. High molecular weight material not linked to the LPS was purified from S. sonnei and confirmed by NMR to contain the specific sugars of the S. sonnei OAg. Deletion of genes homologous to the group 4 capsule synthesis cluster, previously described in Escherichia coli, abolished the generation of the high molecular weight OAg material. This OAg capsule strongly affects the virulence of S. sonnei. Uncapsulated knockout bacteria were highly invasive in vitro and strongly inflammatory in the rabbit intestine. But, the lack of capsule reduced the ability of S. sonnei to resist complement-mediated killing and to spread from the gut to peripheral organs. In contrast, overexpression of the capsule decreased invasiveness in vitro and inflammation in vivo compared to the wild type. In conclusion, the data indicate that in S. sonnei expression of the capsule modulates bacterial pathogenesis resulting in balanced capabilities to invade and persist in the host environment.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: CAM, AS, and CG are employees of Novartis Vaccines Institute for Global Health, Siena, Italy. This does not alter the authors’ adherence to all the PLoS Pathogens policies on sharing data and materials.
Figures
References
-
- Lozano R, Naghavi M, Foreman K, Lim S, Shibuya K, Aboyans V, et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012;380: 2095–2128. 10.1016/S0140-6736(12)61728-0 - DOI - PMC - PubMed
-
- Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet. 2013;382: 209–222. 10.1016/S0140-6736(13)60844-2 - DOI - PubMed
-
- Bennish ML. Potentially lethal complications of shigellosis. Rev Infect Dis. 1991;13 Suppl 4: S319–S324. - PubMed
-
- Struelens MJ, Patte D, Kabir I, Salam A, Nath SK, Butler T. Shigella septicemia: prevalence, presentation, risk factors, and outcome. J Infect Dis. 1985;152: 784–790. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
