

A novel dual kinase function of the RET proto-oncogene negatively regulates activating transcription factor 4-mediated apoptosis
- PMID: 25795775
- PMCID: PMC4416875
- DOI: 10.1074/jbc.M114.619833
A novel dual kinase function of the RET proto-oncogene negatively regulates activating transcription factor 4-mediated apoptosis
Abstract
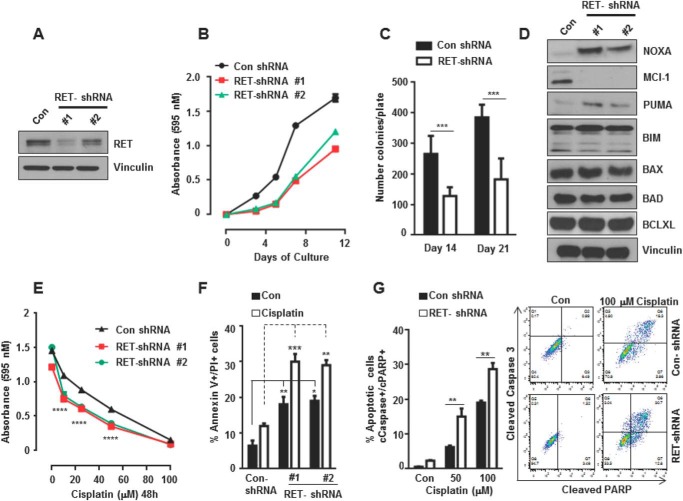
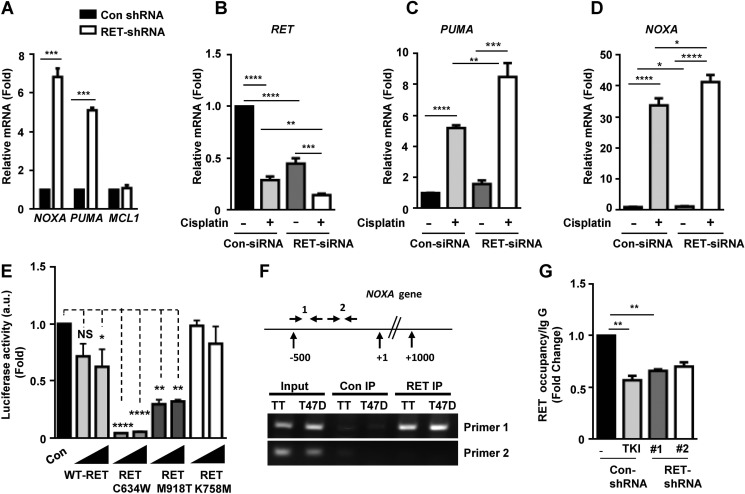
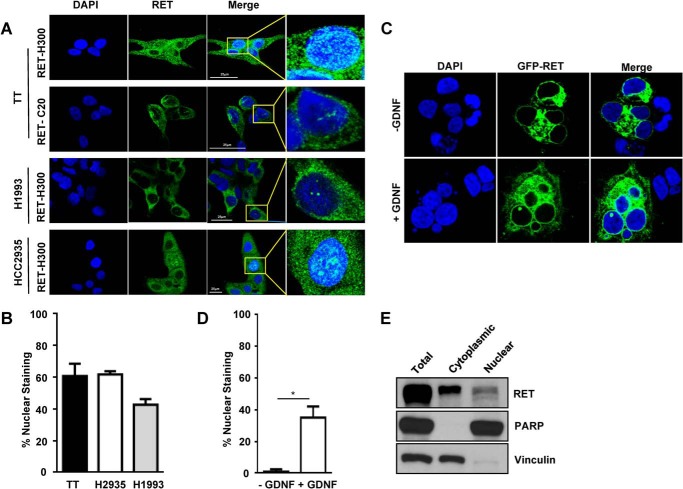
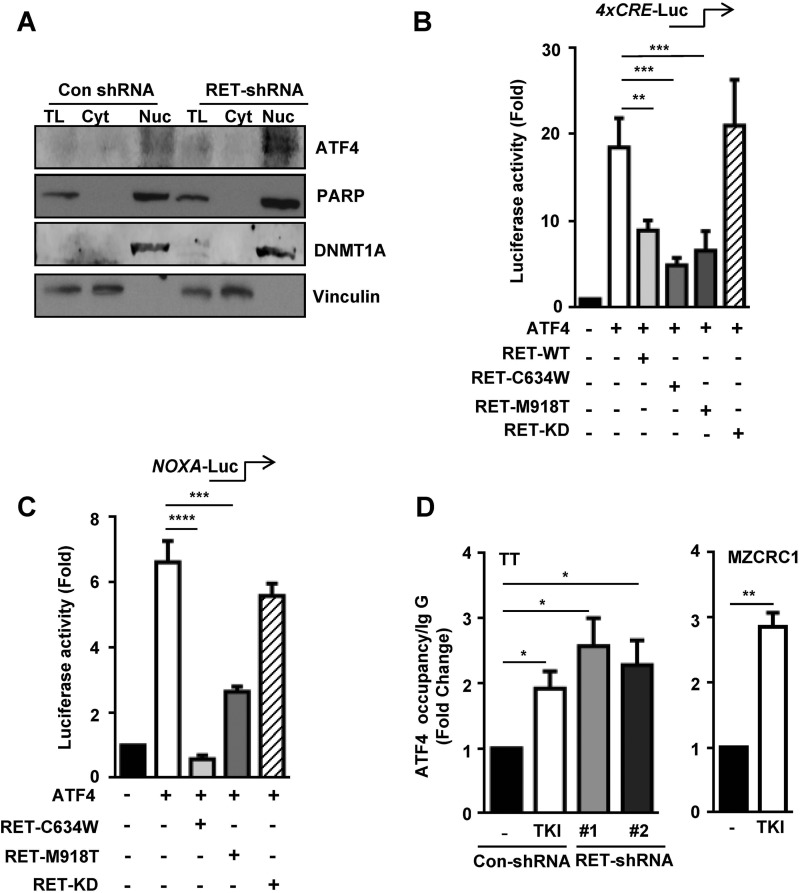
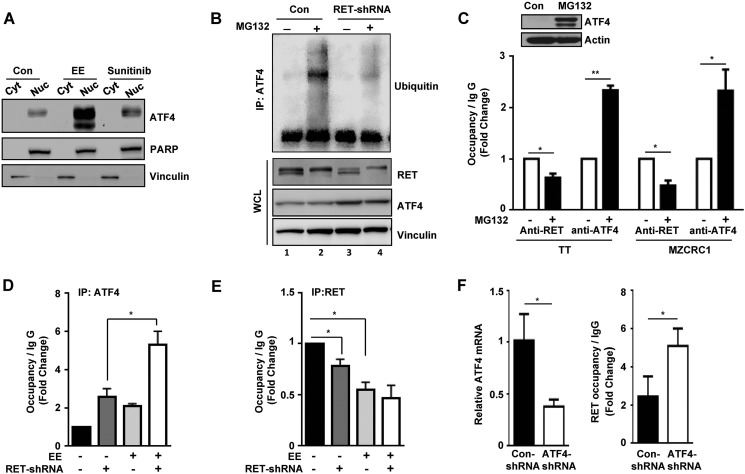
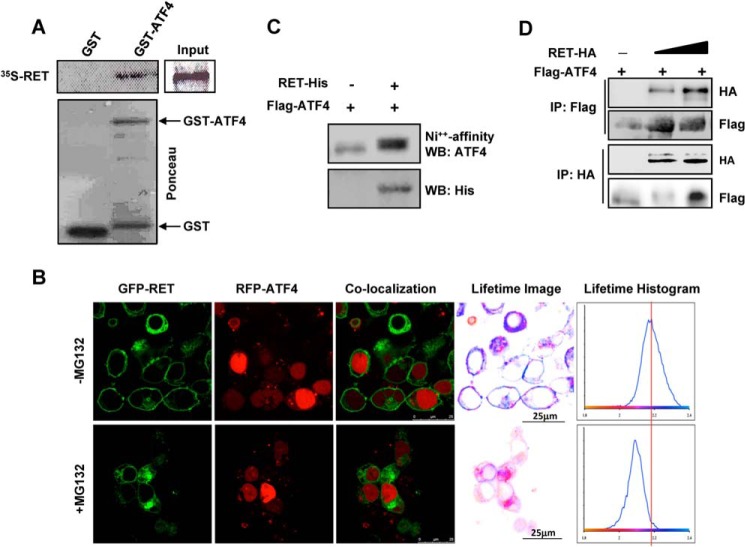
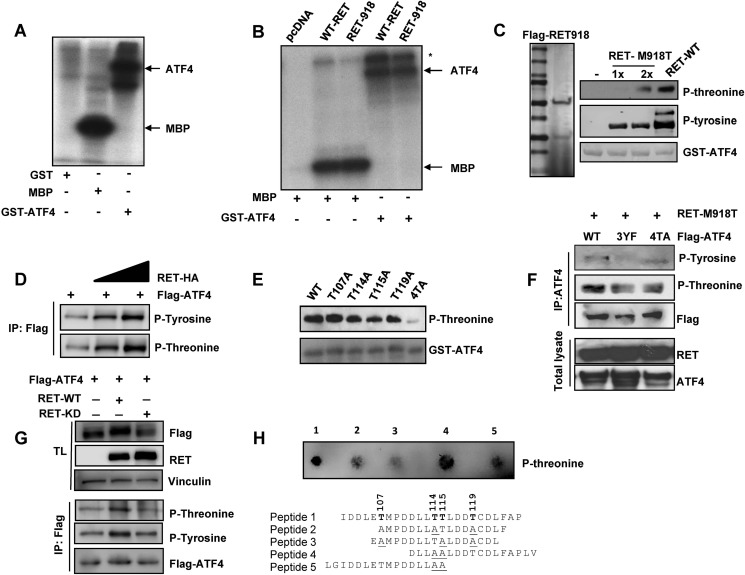
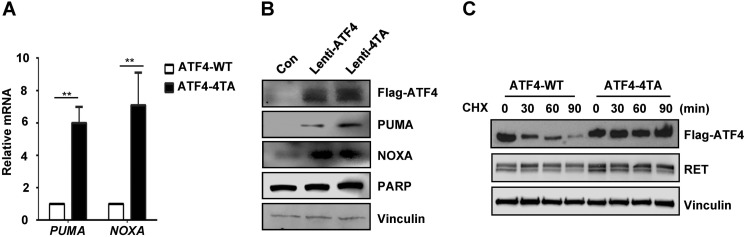
The RET proto-oncogene, a tyrosine kinase receptor, is widely known for its essential role in cell survival. Germ line missense mutations, which give rise to constitutively active oncogenic RET, were found to cause multiple endocrine neoplasia type 2, a dominant inherited cancer syndrome that affects neuroendocrine organs. However, the mechanisms by which RET promotes cell survival and prevents cell death remain elusive. We demonstrate that in addition to cytoplasmic localization, RET is localized in the nucleus and functions as a tyrosine-threonine dual specificity kinase. Knockdown of RET by shRNA in medullary thyroid cancer-derived cells stimulated expression of activating transcription factor 4 (ATF4), a master transcription factor for stress-induced apoptosis, through activation of its target proapoptotic genes NOXA and PUMA. RET knockdown also increased sensitivity to cisplatin-induced apoptosis. We observed that RET physically interacted with and phosphorylated ATF4 at tyrosine and threonine residues. Indeed, RET kinase activity was required to inhibit the ATF4-dependent activation of the NOXA gene because the site-specific substitution mutations that block threonine phosphorylation increased ATF4 stability and activated its targets NOXA and PUMA. Moreover, chromatin immunoprecipitation assays revealed that ATF4 occupancy increased at the NOXA promoter in TT cells treated with tyrosine kinase inhibitors or the ATF4 inducer eeyarestatin as well as in RET-depleted TT cells. Together these findings reveal RET as a novel dual kinase with nuclear localization and provide mechanisms by which RET represses the proapoptotic genes through direct interaction with and phosphorylation-dependent inactivation of ATF4 during the pathogenesis of medullary thyroid cancer.
Keywords: Apoptosis; Thyroid; Transcription Regulation; Transcription Repressor; Tyrosine-Protein Kinase (Tyrosine Kinase); Ubiquitylation (Ubiquitination).
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Wells S. A., Jr., Santoro M. (2009) Targeting the RET pathway in thyroid cancer. Clin. Cancer Res. 15, 7119–7123 - PubMed
-
- Zeng Q., Cheng Y., Zhu Q., Yu Z., Wu X., Huang K., Zhou M., Han S., Zhang Q. (2008) The relationship between overexpression of glial cell-derived neurotrophic factor and its RET receptor with progression and prognosis of human pancreatic cancer. J. Int. Med. Res. 36, 656–664 - PubMed
-
- Díaz-Beyá M., Navarro A., Ferrer G., Díaz T., Gel B., Camós M., Pratcorona M., Torrebadell M., Rozman M., Colomer D., Monzo M., Esteve J. (2013) Acute myeloid leukemia with translocation (8;16)(p11;p13) and MYST3-CREBBP rearrangement harbors a distinctive microRNA signature targeting RET proto-oncogene. Leukemia 27, 595–603 - PubMed
-
- Nikolsky Y., Sviridov E., Yao J., Dosymbekov D., Ustyansky V., Kaznacheev V., Dezso Z., Mulvey L., Macconaill L. E., Winckler W., Serebryiskaya T., Nikolskaya T., Polyak K. (2008) Genome-wide functional synergy between amplified and mutated genes in human breast cancer. Cancer Res. 68, 9532–9540 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
