

An 18 kDa scaffold protein is critical for Staphylococcus epidermidis biofilm formation
- PMID: 25799153
- PMCID: PMC4370877
- DOI: 10.1371/journal.ppat.1004735
An 18 kDa scaffold protein is critical for Staphylococcus epidermidis biofilm formation
Abstract
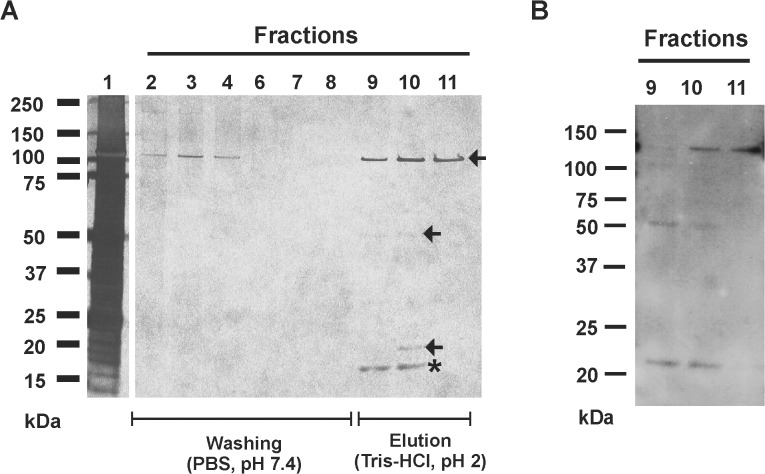
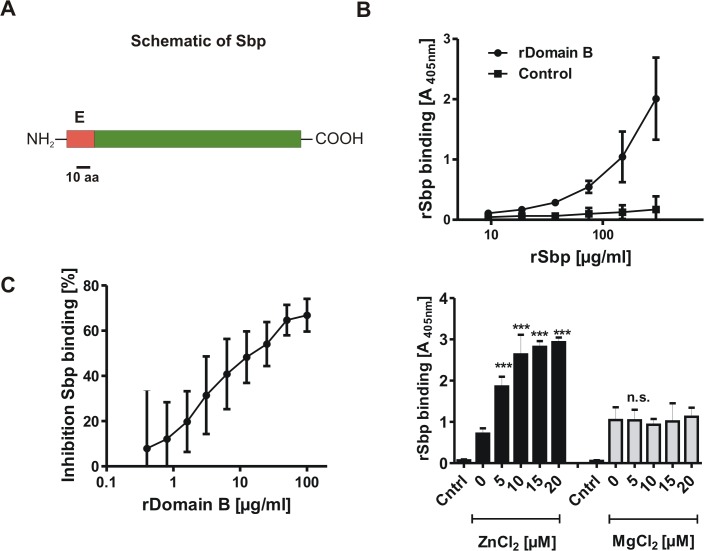
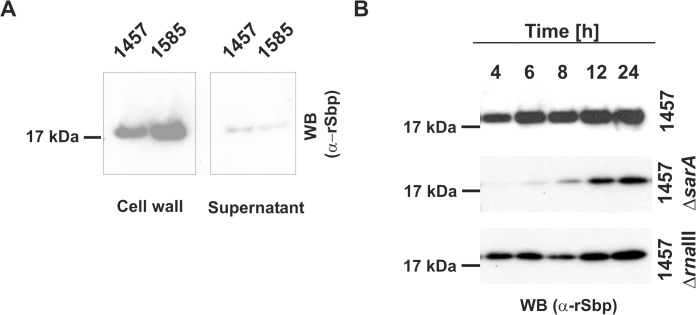
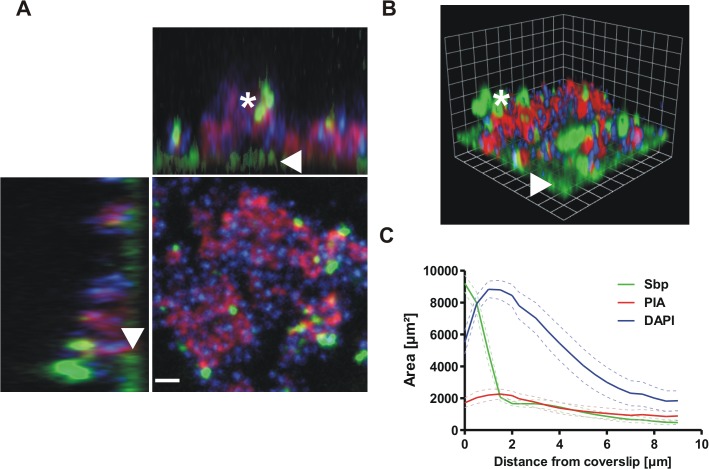
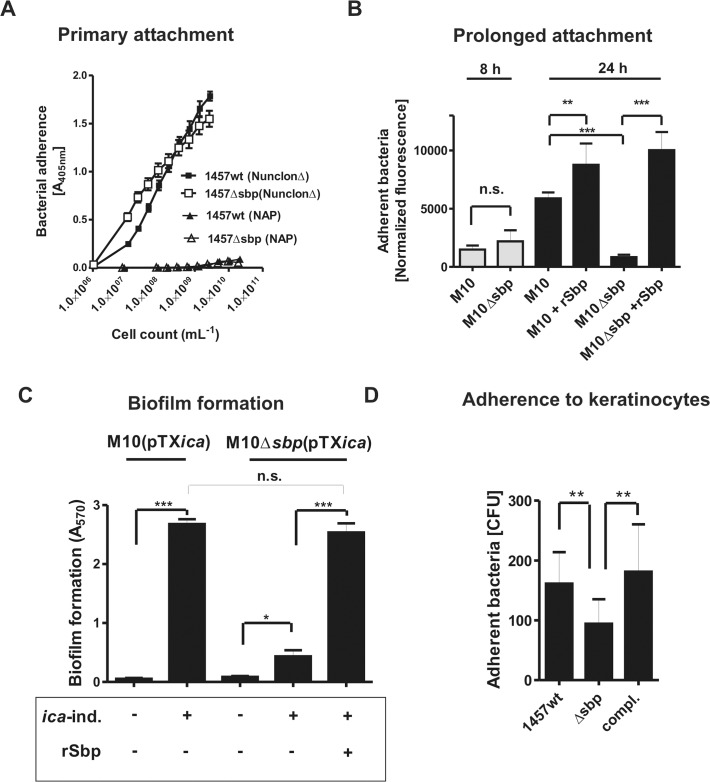
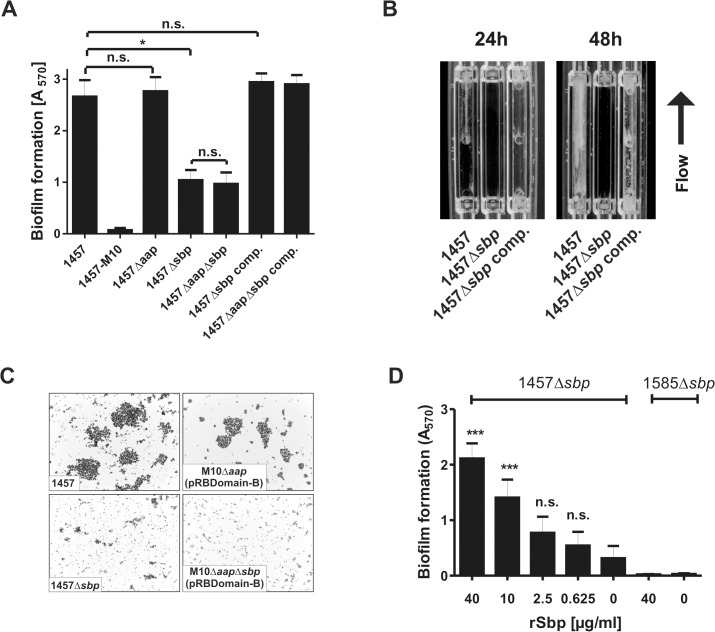
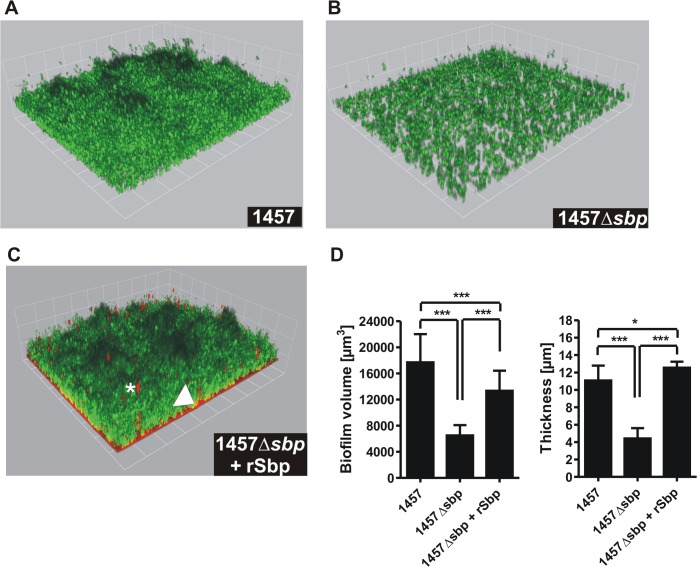
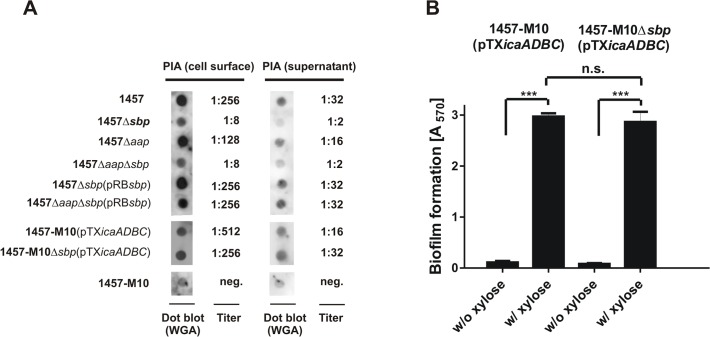
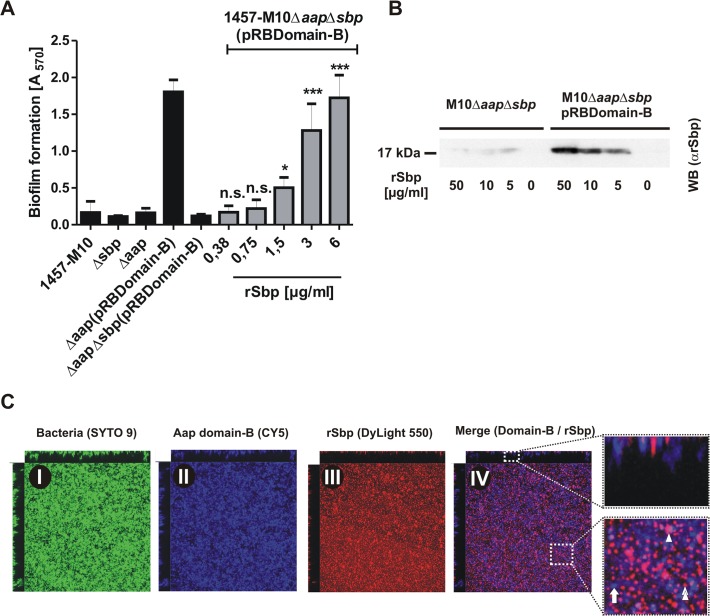
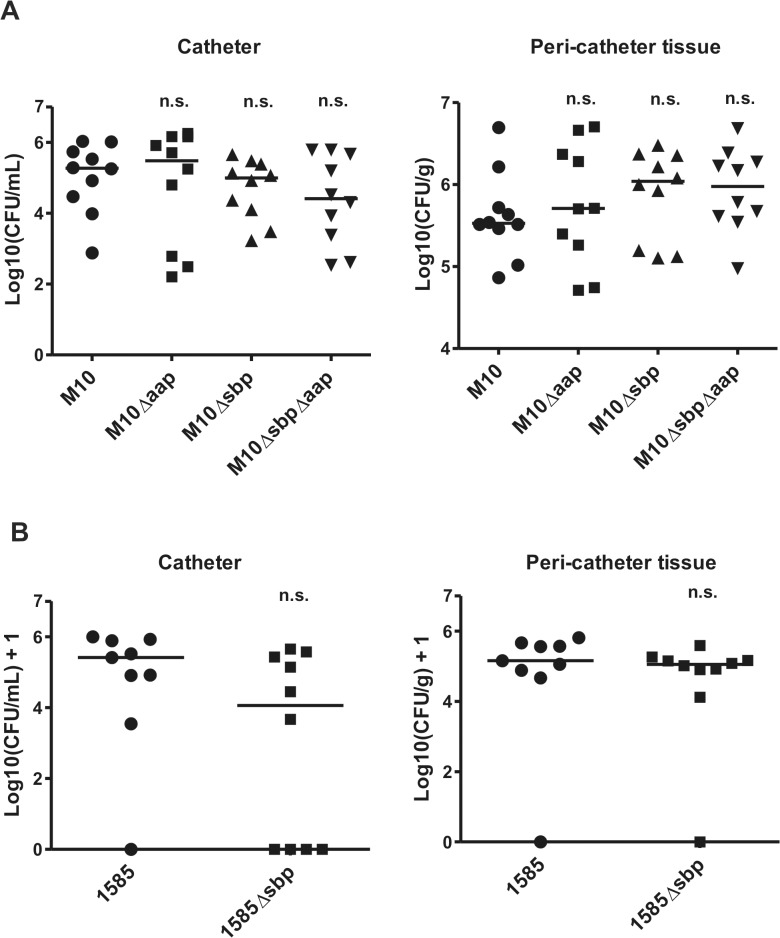
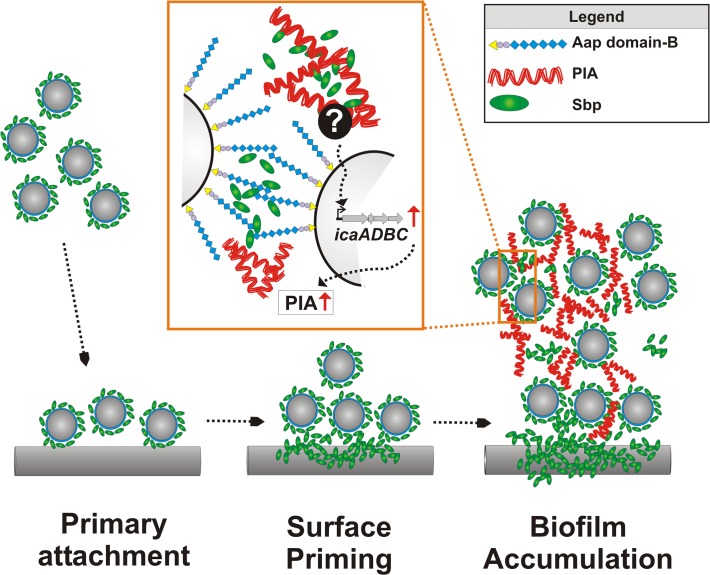
Virulence of the nosocomial pathogen Staphylococcus epidermidis is crucially linked to formation of adherent biofilms on artificial surfaces. Biofilm assembly is significantly fostered by production of a bacteria derived extracellular matrix. However, the matrix composition, spatial organization, and relevance of specific molecular interactions for integration of bacterial cells into the multilayered biofilm community are not fully understood. Here we report on the function of novel 18 kDa Small basic protein (Sbp) that was isolated from S. epidermidis biofilm matrix preparations by an affinity chromatographic approach. Sbp accumulates within the biofilm matrix, being preferentially deposited at the biofilm-substratum interface. Analysis of Sbp-negative S. epidermidis mutants demonstrated the importance of Sbp for sustained colonization of abiotic surfaces, but also epithelial cells. In addition, Sbp promotes assembly of S. epidermidis cell aggregates and establishment of multilayered biofilms by influencing polysaccharide intercellular-adhesin (PIA) and accumulation associated protein (Aap) mediated intercellular aggregation. While inactivation of Sbp indirectly resulted in reduced PIA-synthesis and biofilm formation, Sbp serves as an essential ligand during Aap domain-B mediated biofilm accumulation. Our data support the conclusion that Sbp serves as an S. epidermidis biofilm scaffold protein that significantly contributes to key steps of surface colonization. Sbp-negative S. epidermidis mutants showed no attenuated virulence in a mouse catheter infection model. Nevertheless, the high prevalence of sbp in commensal and invasive S. epidermidis populations suggests that Sbp plays a significant role as a co-factor during both multi-factorial commensal colonization and infection of artificial surfaces.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Wisplinghoff H, Seifert H, Tallent SM, Bischoff T, Wenzel RP, Edmond MB. Nosocomial bloodstream infections in pediatric patients in United States hospitals: epidemiology, clinical features and susceptibilities. Pediatr Infect Dis J 2003. Aug;22(8):686–91. - PubMed
-
- Wisplinghoff H, Seifert H, Wenzel RP, Edmond MB. Current trends in the epidemiology of nosocomial bloodstream infections in patients with hematological malignancies and solid neoplasms in hospitals in the United States. Clin Infect Dis 2003. May 1;36(9):1103–10. - PubMed
-
- Darouiche RO. Treatment of infections associated with surgical implants. N Engl J Med 2004. Apr 1;350(14):1422–9. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
