

Quantitative bead-based flow cytometry for assaying Rab7 GTPase interaction with the Rab-interacting lysosomal protein (RILP) effector protein
- PMID: 25800855
- PMCID: PMC6033261
- DOI: 10.1007/978-1-4939-2569-8_28
Quantitative bead-based flow cytometry for assaying Rab7 GTPase interaction with the Rab-interacting lysosomal protein (RILP) effector protein
Abstract
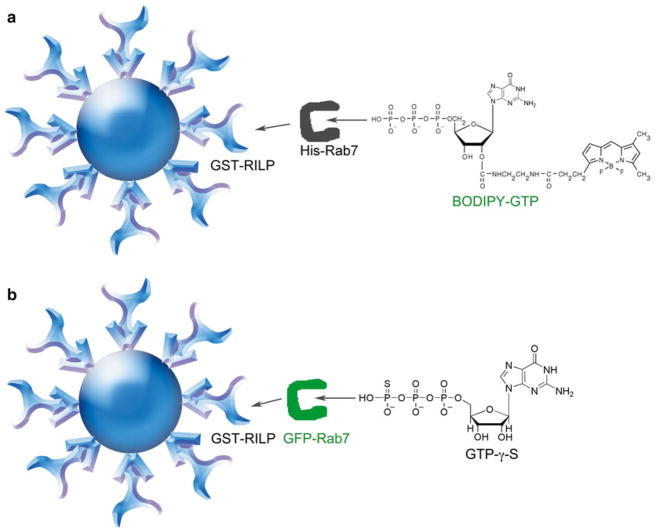
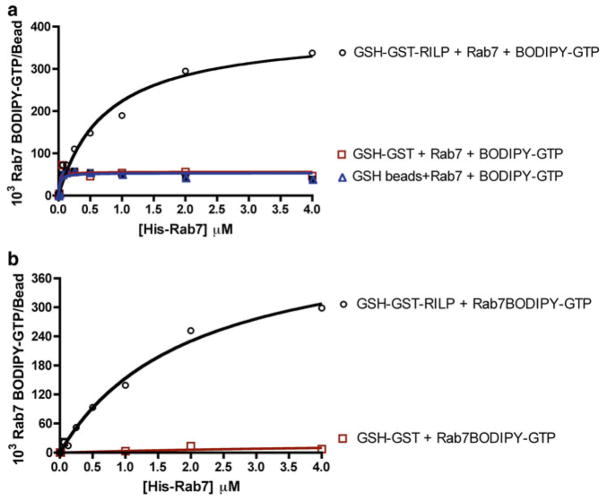
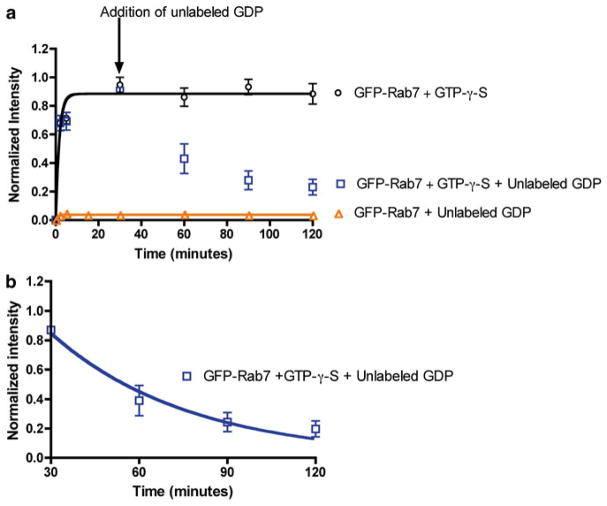
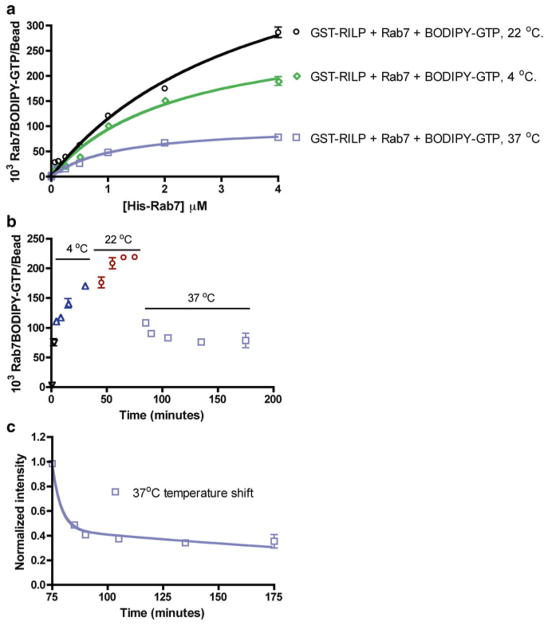
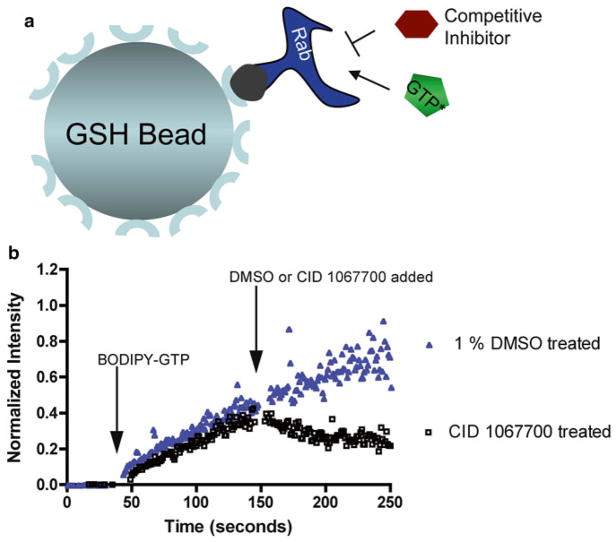
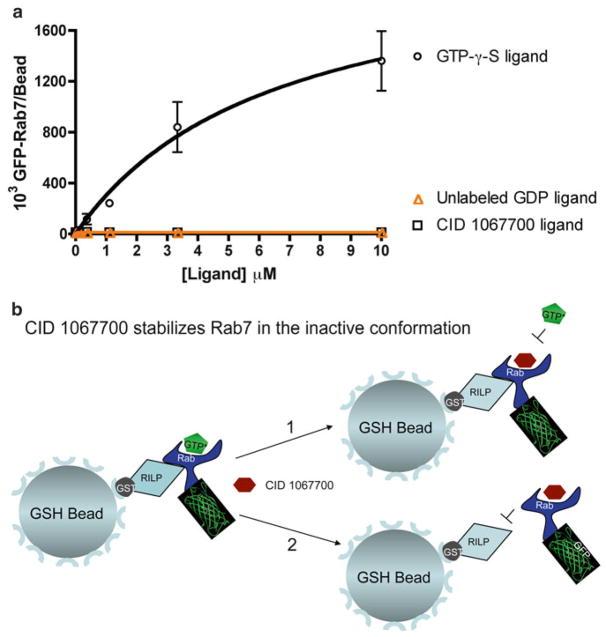
Rab7 facilitates vesicular transport and delivery from early endosomes to late endosomes as well as from late endosomes to lysosomes. The role of Rab7 in vesicular transport is dependent on its interactions with effector proteins, among them Rab-interacting lysosomal protein (RILP), which aids in the recruitment of active Rab7 (GTP-bound) onto dynein-dynactin motor complexes to facilitate late endosomal transport on the cytoskeleton. Here we detail a novel bead-based flow cytometry assay to measure Rab7 interaction with the Rab-interacting lysosomal protein (RILP) effector protein and demonstrate its utility for quantitative assessment and studying drug-target interactions. The specific binding of GTP-bound Rab7 to RILP is readily demonstrated and shown to be dose-dependent and saturable enabling K d and B max determinations. Furthermore, binding is nearly instantaneous and temperature-dependent. In a novel application of the assay method, a competitive small molecule inhibitor of Rab7 nucleotide binding (CID 1067700 or ML282) is shown to inhibit the Rab7-RILP interaction. Thus, the assay is able to distinguish that the small molecule, rather than incurring the active conformation, instead 'locks' the GTPase in the inactive conformation. Together, this work demonstrates the utility of using a flow cytometry assay to quantitatively characterize protein-protein interactions involving small GTPases and which has been adapted to high-throughput screening. Further, the method provides a platform for testing small molecule effects on protein-protein interactions, which can be relevant to drug discovery and development.
Figures
References
-
- Meresse S, Gorvel JP, Chavrier P. The rab7 GTPase resides on a vesicular compartment connected to lysosomes. J Cell Sci. 1995;108:3349–3358. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
