

T cell-extrinsic CD18 attenuates antigen-dependent CD4+ T cell activation in vivo
- PMID: 25801431
- PMCID: PMC4404034
- DOI: 10.4049/jimmunol.1401328
T cell-extrinsic CD18 attenuates antigen-dependent CD4+ T cell activation in vivo
Abstract
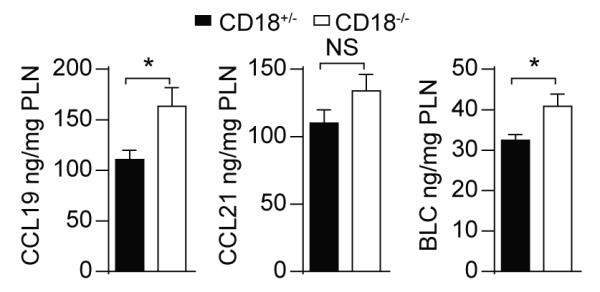
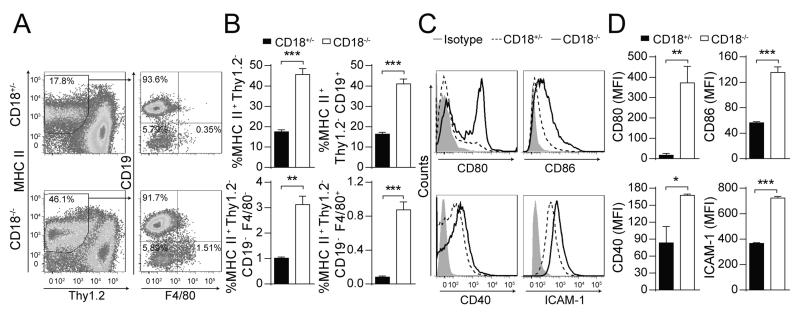
The β2 integrins (CD11/CD18) are heterodimeric leukocyte adhesion molecules expressed on hematopoietic cells. The role of T cell-intrinsic CD18 in trafficking of naive T cells to secondary lymphoid organs and in Ag-dependent T cell activation in vitro and in vivo has been well defined. However, the T cell-extrinsic role for CD18, including on APC, in contributing to T cell activation in vivo is less well understood. We examined the role for T cell-extrinsic CD18 in the activation of wild-type CD4(+) T cells in vivo through the adoptive transfer of DO11.10 Ag-specific CD4(+) T cells into CD18(-/-) mice. We found that T cell-extrinsic CD18 was required for attenuating OVA-induced T cell proliferation in peripheral lymph nodes (PLN). The increased proliferation of wild-type DO11.10 CD4(+) T cells in CD18(-/-) PLN was associated with a higher percentage of APC, and these APC demonstrated an increased activation profile and increased Ag uptake, in particular in F4/80(+) APC. Depletion of F4/80(+) cells both reduced and equalized Ag-dependent T cell proliferation in CD18(-/-) relative to littermate control PLN, demonstrating that these cells play a critical role in the enhanced T cell proliferation in CD18(-/-) mice. Consistently, CD11b blockade, which is expressed on F4/80(+) macrophages, enhanced the proliferation of DO11.10 CD4(+) T cells in CD18(+/-) PLN. Thus, in contrast to the T cell-intrinsic essential role for CD18 in T cell activation, T cell-extrinsic expression of CD18 attenuates Ag-dependent CD4(+) T cell activation in PLN in vivo.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures







References
-
- Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell. 2002;110:673–687. - PubMed
-
- Larson RS, Springer TA. Structure and function of leukocyte integrins. Immunol Rev. 1990;114:180–217. - PubMed
-
- Bunting M, Harris ES, McIntyre TM, Prescott SM, Zimmerman GA. Leukocyte adhesion deficiency syndromes: adhesion and tethering defects involving beta 2 integrins and selectin ligands. Curr Opin Hematol. 2002;9:30–35. - PubMed
-
- Kishimoto TK, Hollander N, Roberts TM, Anderson DC, Springer TA. Heterogeneous mutations in the beta subunit common to the LFA-1, Mac-1, and p150,95 glycoproteins cause leukocyte adhesion deficiency. Cell. 1987;50:193–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
