

Roles of reactive oxygen species-degrading enzymes of Francisella tularensis SCHU S4
- PMID: 25802058
- PMCID: PMC4432764
- DOI: 10.1128/IAI.02488-14
Roles of reactive oxygen species-degrading enzymes of Francisella tularensis SCHU S4
Abstract
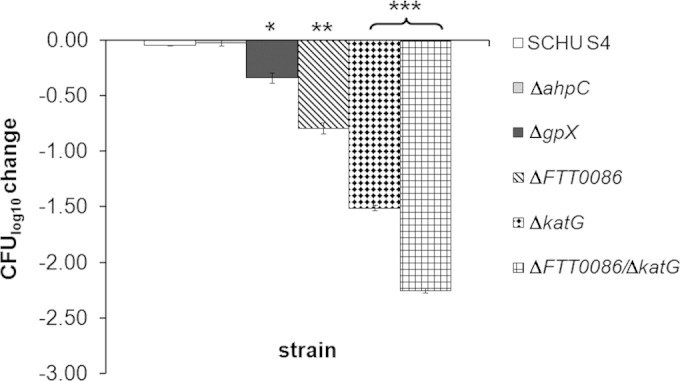
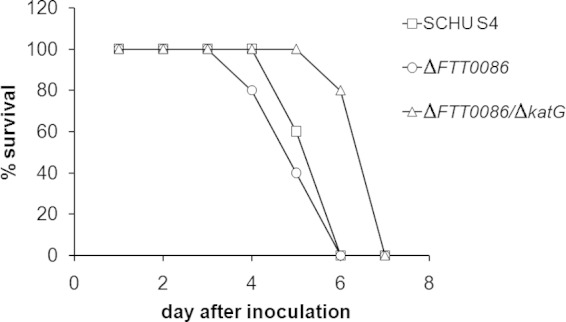
Francisella tularensis is a facultative intracellular bacterium utilizing macrophages as its primary intracellular habitat and is therefore highly capable of resisting the effects of reactive oxygen species (ROS), potent mediators of the bactericidal activity of macrophages. We investigated the roles of enzymes presumed to be important for protection against ROS. Four mutants of the highly virulent SCHU S4 strain with deletions of the genes encoding catalase (katG), glutathione peroxidase (gpx), a DyP-type peroxidase (FTT0086), or double deletion of FTT0086 and katG showed much increased susceptibility to hydrogen peroxide (H2O2) and slightly increased susceptibility to paraquat but not to peroxynitrite (ONOO(-)) and displayed intact intramacrophage replication. Nevertheless, mice infected with the double deletion mutant showed significantly longer survival than SCHU S4-infected mice. Unlike the aforementioned mutants, deletion of the gene coding for alkyl-hydroperoxide reductase subunit C (ahpC) generated a mutant much more susceptible to paraquat and ONOO(-) but not to H2O2. It showed intact replication in J774 cells but impaired replication in bone marrow-derived macrophages and in internal organs of mice. The live vaccine strain, LVS, is more susceptible than virulent strains to ROS-mediated killing and possesses a truncated form of FTT0086. Expression of the SCHU S4 FTT0086 gene rendered LVS more resistant to H2O2, which demonstrates that the SCHU S4 strain possesses additional detoxifying mechanisms. Collectively, the results demonstrate that SCHU S4 ROS-detoxifying enzymes have overlapping functions, and therefore, deletion of one or the other does not critically impair the intracellular replication or virulence, although AhpC appears to have a unique function.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures

References
-
- Dennis DT, Inglesby TV, Henderson DA, Bartlett JG, Ascher MS, Eitzen E, Fine AD, Friedlander AM, Hauer J, Layton M, Lillibridge SR, McDade JE, Osterholm MT, O'Toole T, Parker G, Perl TM, Russell PK, Tonat K. 2001. Tularemia as a biological weapon: medical and public health management. JAMA 285:2763–2773. doi:10.1001/jama.285.21.2763. - DOI - PubMed
-
- Fortier AH, Green SJ, Polsinelli T, Jones TR, Crawford RM, Leiby DA, Elkins KL, Meltzer MS, Nacy CA. 1994. Life and death of an intracellular pathogen: Francisella tularensis and the macrophage. Immunol Ser 60:349–361. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
